Vol. 3° - III.
Altri geni di RIPARTIZIONE DEL NERO E DEL ROSSO
|
|
Oltre ai geni della serie E ne esistono altri che
agiscono sulla ripartizione dell’eumelanina e della feomelanina. Possiamo
distinguere tali geni in due gruppi fondamentali:
· geni che estendono il nero
·
geni che restringono il nero.
1. geni di ESTENSIONE DEL NERO
Oltre agli alleli E e ER, esistono altri fattori che possono intensificare l’effetto
eumelanizzante, capaci cosě di giocare un ruolo importante sia sui colori
primari che secondari. Nonostante la loro importanza, spesso la conoscenza di
questi intensificatori del nero č piuttosto scarsa.
Gli interessi del passato sugli
intensificatori dell’eumelanina si concentrarono su ceppi neri, o quasi, che
non presentavano un piumino giovanile nero né gli effetti dominanti ed
epistatici della mutazione E. Per fare un esempio, la varietŕ Brassy back - schiena d’ottone - del
Combattente inglese antico č omozigote sia per il gene e+ che per un gene autosomico intensificatore dell’eumelanina.
Di tutti i fenotipi descritti, solo la linea Massachusetts
Recessive Black č totalmente nera e si riproduce fedelmente; essa č
caratterizzata da un piumino giovanile come quello presentato da eb
e da un iporachide bianco nei maschi adulti. Quando tale linea viene
incrociata con ceppi e+/e+
e con ceppi eb/eb,
l’effetto piů netto č una dominanza incompleta. Si č quindi ipotizzato
che, a determinare il piumaggio nero non-E, possano
intervenire 3, se non addirittura 4 geni.
1.1. Melanotico
|
Ml
- melanotic |
Autosomico
incompletamente dominante
Gruppo
di associazione III - cromosoma 1
Dagli
studi sul Brassy
back
appena citati si č giunti alla conclusione che in questa varietŕ č
implicato un solo gene eumelanizzante, il melanotico Ml, che č appunto una mutazione
autosomica dotata di dominanza incompleta in grado di estendere l’eumelanina a quelle aree del piumaggio
occupate da feomelanina.
Tutti i soggetti
omozigoti per Ml sono praticamente neri, indipendentemente dall’allele
presente nel locus E. Solo i soggetti frumento risentono in modo meno marcato
dell’omozigosi per Ml. Sono soprattutto le femmine, sia eWh che ey,
a presentare un mantello in cui la colorazione di base č ancora riconoscibile
pur essendo bruno scuro. I maschi corrispondenti si avvicinano invece al nero
esteso. Da ricordare che anche il petto salmone della femmina dorata č poco
sensibile a Ml, conservandosi praticamente intatto o perlomeno evidente.
I soggetti
eterozigoti per Ml sono nettamente piů scuri del normale, specie a
livello della testa e della mantellina. In questa situazione le femmine
frumento sono quelle a risentirne di meno, in quanto presentano talora solo
alcune tracce di nero distribuite irregolarmente sul mantello.
Nel pulcino
il gene del melanismo ha scarsa influenza sul piumino.
Dopo essere stato identificato, si č scoperto che Ml
svolge un ruolo molto importante sia in alcune colorazioni di base, sia in
colorazioni secondarie come quaglia,
orlato semplice e doppio.
Gli sforzi, non seguiti da successo, per determinare la
base genetica di certe linee nero
recessivo, suggeriscono l’esistenza di piů di una mutazione in grado di
intensificare l’eumelanina. Da sola, la mutazione Ml non č in grado di rendere nero un soggetto di tipo selvatico,
in quanto rimangono delle tracce di rosso confinate alle spalle.
Inoltre, i geni melanotici
ipotizzati da diversi ricercatori potrebbero non essere identici. Il gene
eumelanizzante di Crawford ha un legame molto stretto con quello della cresta
a pisello P (0,3% di crossover), mentre lo stesso gene di Carefoot
si trova a 46 unitŕ crossover dal locus occupato dal gene P. Le
relazioni di linkage
tra questi due geni eumelanizzanti e altri due marcatori, Pg e Db, concordano con le
vedute di altri autori. Per cui il gene Ml di Crawford probabilmente
rappresenta una seconda mutazione eumelanizzante localizzata sullo stesso
cromosoma, oppure č una traslocazione di Ml in un locus strettamente legato
al gene P. Naturalmente la relazione tra i 2 geni melanotici deve ancora
essere chiarita.
Si potrebbe affermare che Ml č l’inverso di Db,
in quanto una singola dose di questo secondo gene č in grado non di
estendere, bensě di restringere il nero alla sola mantellina, dipendendo
ovviamente dal genotipo di base.
Il
linkage di Db con P
e Ml, corrispondente a circa 28 e 10 unitŕ rispettivamente,
implica un linkage tra P e Ml di circa 18 oppure
38
unitŕ,
a seconda che il locus Ml si trovi tra P e Db oppure in un altro punto. Il
linkage tra Ml e P di circa 46 unitŕ crossover, come calcolato
da Carefoot, suggerisce che il locus Ml č piů distante da P di quanto non lo
sia Db. Nel 1987 Carefoot ha potuto dimostrare che il locus Ml
risiede tra Db e Pg, da cui deriva che l’ordine dei 3 loci č Db_Ml_Pg, con Pg che si trova all’estremitŕ opposta
rispetto a P, per cui la posizione relativa dei loci č la seguente: P_Db_Ml_Pg. Questi loci appartengono al III
gruppo
di associazione (cromosoma 1) e hanno un linkage di circa 28, 10 e 10 unitŕ
rispettivamente.
Tuttavia, a causa della somiglianza tra i fenotipi
descritti da Crawford nel 1986 e da Carefoot nel 1987,
sono necessari studi ulteriori per giungere a un chiarimento delle
osservazioni dei due ricercatori; non sarebbe strano il fatto che essi
stessero studiando due mutazioni diverse dotate di effetti simili.
Dall’incrocio tra un dorato puro e un soggetto Ml_Ml, si
ottengono soggetti dorato scuro:
| Maschio dorato | Femmina
dorata e melanotica |
||
|
e+ ml+/e+ ml+ |
e+ Ml/e+ Ml |
||
| e+
= dorato ml+ = non estensione melanica, allele di Ml di estensione melanica |
|||
| 100%
dorato scuro le femmine con petto salmonato |
gameti femminili |
||
| e+ Ml | e+ Ml | ||
|
gameti maschili |
e+ ml+ | e+ ml+/e+ Ml | e+ ml+/e+ Ml |
| e+ ml+ | e+ ml+/e+ Ml | e+ ml+/e+ Ml | |
Se si incrociano galli frumento scuro per la presenza di Ml_Ml, si ottengono soggetti
frumento, ma con testa scura, in quanto Ml č presente allo stato eterozigote
dopo l’incrocio:
| Maschio frumento scuro | Femmina frumento | ||
|
ey Ml/ey Ml |
ey ml+/ey ml+ |
||
| ey
= frumento recessivo ml+ = non estensione melanica, allele di Ml di estensione melanica |
|||
| 100% frumento testa scura |
gameti femminili |
||
| ey ml+ | ey ml+ | ||
|
gameti maschili |
ey Ml | ey Ml/ey ml+ | ey Ml/ey ml+ |
| ey Ml | ey Ml/ey ml+ | ey Ml/ey ml+ | |
Vediamo cosa accade se un
soggetto frumento a testa scura
viene a sua volta incrociato con soggetti neri, omozigoti per Ml, ma
eterozigoti per il gene frumento:
| Maschio melanotico eterozigote in E | Femmina
frumento testa scura |
||
|
e+ Ml/ey Ml |
ey Ml/ey ml+ |
||
| ey
= frumento recessivo e+ = dorato ml+ = non estensione melanica, allele di Ml di estensione melanica |
|||
| 25%
melanotico, frumento_dorato 25% frumento_dorato, testa scura 25% frumento scuro 25% frumento testa scura |
gameti femminili |
||
| ey Ml | ey ml+ | ||
|
gameti maschili |
e+ Ml | e+ Ml/ey Ml | e+ Ml/ey ml+ |
| ey Ml | ey Ml/ey Ml | ey Ml/ey ml+ | |
Ml č pure responsabile della melanina epidermica,
che puň essere inibita da B; per cui i tarsi di questi barrati sono sempre
privi di tale pigmento, o presentano solo una lieve sfumatura cenere.
1.2. Ebano
|
Ebano |
Nelle
varietŕ nere a tarsi
gialli, Ml gioca il ruolo maggiore e forse č associato a geni
minori di estensione del nero, detti ebano, alcuni dei quali sono
necessari per completare l’estensione del nero alle spalle.
Sebbene esistano molti esempi di piumaggio nero tra le
razze riportate negli standard, la loro somiglianza č piů fenotipica che
genotipica. Senza dubbio una buona parte si basa sul gene del nero esteso E,
oppure sul gene ER.
Puň esistere tuttavia un piumaggio totalmente nero in assenza di questi due
geni.
Cote nel 1976 dimostrň la presenza di geni
intensificatori dell’eumelanina nei soggetti neri con genotipo E e ER, forse deputati a incrementare gli effetti epistatici di
questi due alleli. Nello studio di Cote nessuno dei soggetti eterozigoti per E
sviluppň un piumaggio nero esteso in assenza di eumelanizzanti. Il gene Ml
era un componente di tutti i genotipi, eccetto che in una popolazione nera
allevata a caso e nel Combattente Inglese Moderno ER.
Ml era unito al gene dell’orlo nero, fatta eccezione per l’Ancona
pomellata che era omozigote per Ml e per il non-orlato.
Dai risultati ottenuti č stato possibile dedurre con
certezza che i
fenotipi nero esteso
abitualmente sono dotati di intensificatori dell’eumelanina
che hanno lo scopo di aiutare gli alleli della serie E nella loro compito di
estensione del nero. L’allele E, anche da solo, č in grado di trasformare
un soggetto dorato in un soggetto nero. Ma solo un’omozigosi per E e l’aggiunta
di Ml in doppia dose permettono di realizzare il nero integrale.
2. geni di RESTRIZIONE DEL NERO
I fattori capaci di ridurre le
aree eumelanizzate determinano un’estensione di quelle feomelaniche,
colorate in rosso, bruno, fulvo e oro. Pertanto la loro azione si svolge sulla
colorazione di base.
Queste mutazioni si comportano come antagonisti delle aree
nere, e per lo piů sono in grado di determinare un fenotipo columbia o simil-columbia,
in cui l’eumelanina č relegata alla mantellina, alle ali, alle piume dei
tarsi e alla coda. Il termine columbia
indica questo modo di distribuirsi del nero in seguito alla Columbian Exposition at the
Chicago World’s Fair del 1893,
quando soggetti Wyandotte appartenenti a questa varietŕ furono esposti per la
prima volta da Briggs, che aveva giŕ introdotto la Wyandotte bianca nell’universo
avicolo.

Fig. III. 1 - Wyandotte bianco columbia
I geni di restrizione tipo
columbia sono particolarmente efficaci sul mantello delle femmine
non-E e sul petto nero dei maschi dorati.
I geni meglio studiati sono il columbia - Co - il dark brown o marrone scuro -
Db - il mogano - Mh -, mentre altri geni, come quello della diluizione Di e del
piumino chiaro Li sono stati descritti solo sommariamente. Essi interagiscono
con gli alleli della serie E, con altri geni non attivi sul piumaggio (taluni
ipotizzano un’azione favorevole sul peso da parte di Co), forse con altri
geni ancora sconosciuti. Ne scaturiscono i vari fenotipi che descriveremo tra
poco. Inoltre, due fattori di restrizione del nero - Co e Db - giocano un ruolo
importante nei genotipi che sono alla base di molti disegni del piumaggio.
|
Geni
coinvolti nella restrizione dell’eumelanina a carico di mantellina,
ali, tarsi e coda |
||||
| Simbolo | Nome | Dominanza | Effetto sulla feomelanina (1) | Effetto sul nero esteso E |
| Co | Columbia | incompleta | arancio oro | scarso |
| Mh | Mogano | incompleta | scurente | scarso |
| Db | Dark brown (2) | incompleta (3) | arancio marrone |
rende rossastro il
pulcino nero |
| Di | Diluizione | incompleta | diluizione spiccata |
ignoto |
|
1
- effetto
riferito al rosso di tipo selvatico o al color salmone |
||||
Gli effetti di questi geni
risultano
maggiormente evidenti
quando il soggetto ha un genotipo e+ oppure
eb.
Gli effetti sui soggetti E, oppure ER,
sono molto scarsi oppure nulli.
Talune classificazioni includono in questo gruppo anche il
gene della diluizione Di, che invece tratteremo all’inizio della V sezione.
Potremmo discutere all’infinito, perché anche Bl - il gene del blu -
diluisce il nero e lo restringe a tal punto da farlo scomparire negli
omozigoti, S puň diluire gli effetti del suo allele nei soggetti con piumaggio non da
gallina, il gene B - barrato legato al sesso - restringe il nero in barre. Da
questo si arguisce che in biologia le classificazioni sono spesso artificiose.
Facciamo di necessitŕ virtů, accontentandoci dello schematismo che viene per
lo piů proposto. Tra qualche decennio nuove scoperte genetiche sovvertiranno
qualsiasi dogma classificativo.
I
geni di restrizione del nero sono tre:
Co
- columbia
Db
- dark brown - marrone scuro
Mh
- mogano
Sono tutti autosomici e dominanti incompleti.
Mancano ovviamente nella colorazione selvatica, che possiede gli alleli niente
columbia, niente dark brown, niente mogano.
Il combinarsi di queste mutazioni ci offre alcune varietŕ
di piumaggio che hanno come base il columbia, e che fondamentalmente sono tre:
Columbia
propriamente detto
Sia nel maschio che nella
femmina il nero viene circoscritto al collo, alle ali, alla coda, alle piume
dei tarsi, nonché alla sella del gallo (Sussex, Brahma, Wyandotte).
Rosso
coda nera
Sia nel maschio che nella
femmina troviamo scarsa quantitŕ di nero su collo, ali, coda; č presente
poco nero anche sulla sella del gallo (Rhode Island Red e New Hampshire).
Fulvo
Espressione
estrema della restrizione: assenza di nero su collo, ali, coda, sella e tarsi
(Cocincina, Orpington).

Fig. III. 2 - Brahma bianco columbia
2.1. Columbia
|
Co - columbia |
Autosomico
incompletamente dominante
Gruppo
di associazione III - cromosoma 1
Per
dare un’idea delle capacitŕ di questo gene, si pensi che č in grado di
cambiare una Brahma dark, cioč
argento pluriorlata, nella varietŕ che definiamo bianco
columbia, e il maschio tipo Bankiva in un soggetto fulvo columbia. Allo stato eterozigote il gene Co possiede un’azione
completamente dominante nel maschio, ma lo stato omozigote č essenziale nelle femmine
per togliere il nero dal cuscino e dalla parte bassa del petto. Le differenze
nel colore dell’iporachide tra Brahma e Sussex č dovuta al diverso gene di
base, eb - ey
rispettivamente, per cui la restrizione columbia č solo apparentemente
identica. Le femmine Co_Co sono identiche ai loro maschi, fatta eccezione per
la sella, dove esse non presentano il nero. Si puň pertanto sentenziare che
Co č un gene capace di uniformare il piumaggio nei due sessi.
Inizialmente questa mutazione fu classificata come gene
ipostatico in una linea di soggetti bianco recessivo e fu studiata per la
prima volta nella Brahma fulva. Probabilmente č identica alla mutazione ginger
- Gr
[1]
- isolata nella Minorca fulva; tuttavia l'identitŕ non č stata dimostrata con test di
genetica.
I
fenotipi adulti si somigliano quando č in causa uno qualunque dei
seguenti geni: e+,
eb, eWh,
ey. I soggetti omozigoti per il columbia e per la
colorazione di base eb/eb presentano una distribuzione dell’eumelanina come
descritto dall’American Standard of Perfection.
|
Fig.
III. 3
-
Piumaggio bianco columbia |
Gallo
Gallina |
I maschi e+/e+ hanno il classico fenotipo del columbia; le femmine sono
simili a quelle con genotipo eb,
ma presentano alcune punteggiature nere limitate al dorso.
I soggetti eWh/eWh differiscono dal columbia voluto dallo standard
per la scarsa eumelanizzazione, particolarmente evidente a carico delle
lanceolate d’ambo i sessi.
Lo
stato eterozigote per Co č discretamente attivo sul piumaggio dei
maschi in presenza di uno dei suddetti genotipi, anche se residuano tracce di
nero alla sella; nella femmina eterozigote persiste del nero a livello del
cuscino e della parte bassa del petto.
Oltre ad agire sull’eumelanina, Co impartisce alla feomelanina un colore arancio-oro,
o ginger.
Un singolo allele Co č in grado di cambiare in arancio-oro il petto salmone
della femmina selvatica e il mantello frumento.
Non esistono dati indicativi per
ritenere che Co abbia qualche effetto degno di nota sul nero esteso E.
L’espressione
di Co a livello del piumino
č estremamente variabile e dipende dalla presenza di altri geni. In generale,
l’azione sul pulcino risulta dall’interazione tra Co, gli alleli del locus
E e gli intensificatori dell’eumelanina non ancora definiti. In assenza di
questi ultimi e in presenza del gene eb,
il colore del piumino del dorso č grigio chiaro oppure cannella, a seconda
che il pulcino sia argento oppure oro. Quando il colore di fondo č
determinato da e+/e+,
il piumino č simile al precedente, salvo la striscia dorsale che č piů
evidente; in alcuni casi la striscia del capo č appariscente. In presenza del
frumento omozigote, la schiena č essenzialmente chiara, bianca o oro.
Nei soggetti eb
l’aggiunta di intensificatori dell’eumelanina provoca un piumino al dorso
che varia da grigio scuro a nero. Quando l’espressione melanica č al
massimo, persino il ventre č grigio, e l’unica area non nera č la parte
anteriore della faccia. I pulcini piů intensamente eumelanizzati sono
abitualmente femmine, che possono essere facilmente confuse con pulcini
eterozigoti per E. La versione eumelanizzata del piumino selvatico mostra, in
modo tipico, residui delle bande alla schiena e alla testa; tuttavia l’effetto
sul piumino frumento dominante si limita a uno scarso scurimento del dorso.
Co possiede una duplice
influenza:
· modifica la ripartizione del nero relegandolo nei distretti occupati da piume di copertura: mantellina, coda, ali, tarsi
· modifica la ripartizione del nero nel contesto della piuma, per cui, per fare l’esempio del bianco columbia,
nella mantellina il nero si dispone ai due lati del rachide e viene bordato di
bianco, le remiganti hanno il bordo bianco da un solo lato, l’iporachide
nella Brahma č grigio ardesia, talora bluastro, mentre č bianco nella
Sussex.
Attenzione: gli anglofoni denominano buff columbia il nostro columbia o fulvo
columbia, mentre
definiscono columbia
oppure light
il nostro ermellinato o bianco
columbia
[2]
.
Possediamo
tutti gli elementi per fissare i punti chiave dell’azione di Co:
GALLO: quando Co č omozigote in un genotipo e+, eb,
eWh, ey,
otteniamo sempre un fulvo columbia. Quando č in causa il frumento, si ha una
minor quantitŕ di nero sulla sella e sulla mantellina rispetto a quando sono
presenti e+ oppure eb. Quando č presente anche il gene S, allora si ottiene il
bianco columbia ![]() . I galli eterozigoti per Co
hanno praticamente un fenotipo columbia, e raramente il petto č macchiato.
. I galli eterozigoti per Co
hanno praticamente un fenotipo columbia, e raramente il petto č macchiato.
GALLINA: l’omozigosi per Co, su uno
sfondo e+ oppure eb, determina il fulvo columbia e il petto salmonato viene
soppresso. Č per questo che le femmine bianco columbia derivate da e+ hanno un petto bianco. Infatti Co_Co elimina il color
salmone e il gene S determina il bianco. Se Co č eterozigote, esistono
differenze a seconda del genotipo di base:
eb_Co/eb_co+:
una gallina eb possiede del nero nella parte
bassa del petto e al cuscino della coda.
e+_Co/e+_co+:
una gallina con sfondo e+
ha un petto che va dal salmonato all’aranciato e una schiena con pepatura.
ey_Co/ey_co+ eWh_Co/eWh_co+: le femmine frumento sono macchiate alla schiena.
I soggetti frumento sono meno
eumelanizzati nelle aree di restrizione; la feomelanina diventa un arancio-oro,
come accade per la New Hampshire; Co č in grado di accentuare le differenze
tra l’azione dell’argento e dell’oro: questo fatto č di grande aiuto
nel sessaggio dei pulcini S/s+ e s+/W;
sempre il gene columbia intensifica l’espressione del barrato legato al
sesso che si esprime su uno sfondo e+
oppure eb.
2.2. Mogano
|
Mh - mahogany |
Autosomico
incompletamente dominante
Gruppo
di associazione sconosciuto
Manifesta
bene i suoi effetti soprattutto su uno sfondo eb
oppure e+. Oltre a determinare la particolare colorazione
mogano, possiede la capacitŕ di indurre in ambedue i sessi una restrizione
del nero presente al petto, al collo, alla schiena e nelle piccole copritrici
alari. La coda diventa bruno-nero. Anche questo
gene fu isolato per la prima volta nella Minorca fulva. Nel maschio adulto
restringe il nero del petto in modo caratteristico, in quanto sospinge il nero
dall’iporachide verso la parte distale e apicale della piuma, con un effetto dose dipendente
a seconda dell’omo/eterozigosi, per cui gli omozigoti sembrano possedere un disegno pagliettato
di entitŕ variabile, cui fa da sfondo il rosso mogano.
Infatti Mh č stato introdotto nell’Amburgo pagliettata oro
per
l’effetto che dimostra nel favorire la paillette.![]() -
- ![]() .
.
L’azione
di Mh puň essere riassunta
dicendo che in ambedue i sessi causa una riduzione dell’eumelanina a livello
del dorso e delle ali, sospingendo inoltre il nero di ogni singola piuma verso
l’apice.
Un effetto addizionale del mogano č quello di scurire la feomelanina
e con certe combinazioni geniche ne risulta un piumaggio rosso scuro. Pare non
agisca sul colore del piumino. Osservazioni di Warren, confermate da Byerly
& Quinn, hanno messo in evidenza, nei neonati Rhode Island red a cresta
semplice, un dicromatismo del piumino ereditario: l’85% delle
femmine presenta quote di nero variabili da una chiazzetta posta alla base
della testa per giungere a strisce nere su testa, collo e dorso; il 78% dei
maschi non presenta alcuna macchia, piccola o grande che sia.
Anche se Mh non č legato al
sesso, i galli conservano sempre piů nero delle galline. Si rammenta che
esiste un antagonismo tra una maturitŕ
sessuale precoce e una colorazione rossa piů intensa:
i tardoni sono piů scuri. Si č anche potuto concludere che un piumino piů
scuro alla nascita sarebbe predittivo di un piumaggio adulto maggiormente
pigmentato. Il fenotipo rosso coda nera
č proprio della Rhode Island red, la cui feomelanina appartiene a una
varietŕ particolare dimostrata con la fluorescenza. Questa razza possiede
come gene di base ey.

Fig. III. 4 - Rhode Island red
2.3. Marrone scuro
|
Db
- dark brown |
Autosomico
incompletamente dominante
Gruppo
di associazione III - cromosoma 1
Il
nome attribuito a questo gene puň trarre in inganno, in quanto induce a
pensare che questa mutazione sia la causa del marrone scuro osservabile in una
Livorno perniciata scura, mentre in questo caso potrebbe essere implicato un
allele di un altro locus, del locus Dk.
Si č visto che il gene Db, isolato per la prima volta in
pulcini Fayoumi
[3]
con barratura autosomica, trasforma il piumino nero associato al gene ER in un piumino marrone scuro. Il termine dark
brown č ingannevole in quanto si č visto che questa mutazione, isolata e
ricombinata con e+
e con eb, possiede
un effetto non tanto sul piumino del pulcino, quanto sulla colorazione di
base, che si traduce in una distribuzione dell’eumelanina secondo il tipo
columbia.
Gli anglofoni scrivono Fayoumi anche Fayumi. Un egiziano verace mi ha scritto perentoriamente di suo pugno che la traslitterazione della cittŕ dalla quale la razza ha preso il nome č El Faiuom. L’enciclopedia De Agostini su CD riporta El Faiyűm, l’antica Crocodilňpolis, attualmente con 167.000 abitanti e capoluogo dell’omonimo governatorato, 88 km a SSW del Cairo. Situata in una fertile oasi alla sinistra del Nilo, nell'omonima depressione, č nodo stradale e ferroviario e attivo mercato agricolo con industrie tessili (cotonifici, lanifici), della concia e del tabacco. La depressione di El Faiyűm č una regione di circa 1.800 km˛ situata sotto il livello del mare, sulla sinistra del basso corso del Nilo, da cui la separa una stretta striscia desertica alta fino a 157 m sm. A NW il fondo della depressione č occupato dal lago Qârűn le cui acque si trovano a – 44 m. Irrigata dal canale Yűsef, la depressione č coltivata a cereali, cotone, canna da zucchero, vite e olivo. Si tratta di una tra le piů importanti stazioni preistoriche egiziane, sulle rive di un antico bacino lacustre oggi occupato in parte dal lago Qârűn, dove si sono succeduti insediamenti preistorici con datazioni comprese tra circa 6.300 e 5.800 anni fa. Grano, orzo e lino sono comunemente attestati insieme a strumenti per l'agricoltura (falcetti, macine e pestelli). L'economia era integrata dalla pesca e dalla caccia. Oltre ad accette e punte di freccia triangolari, č documentata una ceramica a superficie rossa o scura. Sono stati inoltre rinvenuti molti resti fossili di numerose specie risalenti all'Eocene e all'Oligocene. Famosi sono i Ritratti di Faiyűm: dipinti a encausto o a tempera su tavolette di legno, furono rinvenuti tra le bende delle mummie nelle sepolture dell'oasi del Faiyűm e anche in altre zone lungo il Nilo. L'usanza di fissare il ritratto del defunto sulla mummia č di ascendenza faraonica, ma i ritratti del Faiyűm sono di epoca romana (sec. I-IV dC). In essi, alcuni di autentico valore artistico, altri di modesto livello artigianale, lo stile derivato alla regione da lontane ereditŕ egizie si unisce allo spirito dell'ellenismo e alle piů vivaci correnti figurative dell'arte romana. Encausto: antica tecnica pittorica, in uso giŕ presso i Greci per dipingere su muro, legno, terracotta, marmo o avorio, in cui i colori sono sciolti nella cera fusa per renderli resistenti alla luce e all'acqua, quindi applicati a pennello o a ferro riscaldato. La tecnica del procedimento non č ancor oggi ben chiarita.
Vale la pena ricordare quanto detto a proposito del colore
di base ebc:
il gene Db allo stato omozigote inibisce la curvatura del disegno, per cui lo
spruzzato si trasforma in barrato. Pertanto, la presenza singola o duplice del
gene Db č in grado di modificare la disposizione dell’eumelanina.
Nel gallo tipo Bankiva l’effetto maggiore di Db
č quello di restringere il nero quasi interamente alle ali e alla coda.
Otterremo cosě un fenotipo rosso
coda nera.
Il gene Db fu studiato da Campo & Alvarez nel 1988
in
una razza spagnola detta Villafranquina.
Quando i maschi Villafranquina venivano incrociati con linee-prova dotate del
gene ebc oppure eb, i discendenti avevano un piumaggio simile alla razza
Villafranquina, per cui si dedusse che questa razza contiene l’allele Db
insieme a uno dei due alleli della serie E, anche se č difficile distinguere
quale dei due sia presente.
I
maschi adulti omozigoti per Db, con sfondo oro oppure argento, hanno la
coda nera, le lanceolate della mantellina non sono nere e l’eumelanina di ali e coda č presente in quantitŕ piuttosto ridotta.
Le
femmine adulte omozigoti hanno un disegno che, per quanto riguarda il nero,
somiglia a una distribuzione modificata del tipo columbia, ma che nei soggetti
eb/eb
differisce per la presenza di un simil barrato autosomico nella parte piů
bassa del petto e alla schiena. Anche nelle femmine e+/e+ gli effetti sono simili, solo che Db ha un effetto
relativamente scarso sul salmone del petto e sul mantello delle femmine
frumento.

Fig. III. 5 - Fayoumi argento
Le
femmine eterozigoti somigliano alle omozigoti, eccezion fatta per il
disegno che č piů accentuato.
I
maschi Db/db+
hanno un fenotipo molto variabile. Essendo piů comunemente in causa e+, eb,
eWh, aggiungono una punta rossa alle piume nere del
petto e posseggono un po’ piů di nero alla sella.
Questo gene modifica
anche il colore della feomelanina, col risultato di un piumaggio
arancio-marrone, oppure arancio-bruciacchiato.
Tutto ciň non si verifica sul petto salmone o sul mantello frumento, dove si
nota solo un salmone chiaro. I maschi omozigoti e+,
eb, eWh,
mostrano tutti quanti un leggero ma consistente affievolimento della
feomelanina del dorso, dovuto a un’assenza o a una marcata diluizione di
tale pigmento sui margini distali delle piume della sella.
Tutti
i piumini dovuti agli alleli del locus E vengono modificati da Db,
particolarmente quando sono omozigoti per questo gene.
Gli effetti su E e su ER
si esprimono attraverso un piumino che, anziché essere nero, si fa marrone
scuro e che varia da un marrone scuro nella parte anteriore della faccia sino a
dare un pulcino interamente marrone, e ciň si verifica a seconda della dose
di Db e del restante genotipo.
Quantunque Db si comporti come dominante, si č visto che in presenza di ER agisce come
gene influenzato dal sesso,
essendo dominante nei maschi e recessivo nelle femmine.
In presenza di e+
o di eb uno stato omozigote per Db si traduce in una spiccata
riduzione delle strisce marroni al dorso, che giacciono su di uno sfondo crema
chiaro. Gli eterozigoti sono denunciati da strisce bianco giallastre
interposte piů ampie, e nei soggetti eb
sono caratteristiche le aree marrone chiaro nella parte anteriore della faccia
e nelle aree adiacenti alla parte anteriore della cresta.
In una situazione ebc/ebc induce solo alcune macchioline e una vaga striscia
laterale al dorso su sfondo crema.
Db non si esprime sul piumino frumento.
Quando un pulcino ha un piumino marrone non vuol dire che
Db sia per forza presente.
La
presenza contemporanea di Co e Db
determina il cosiddetto effetto
decolorante, specie nelle aree di restrizione condivise dai due geni.
Questo risultato era attribuito a un altro gene, e recenti studi confermano la
prima affermazione, basandosi sul fatto che tale effetto mostra un linkage col
gene P della cresta a pisello corrispondente a 28 unitŕ
crossover, per cui č determinato da qualche gene presente sul cromosoma 1.
Questo gene č null’altro che l’azione combinata di Co e Db. Per maggiori
dettagli si veda la sezione V.8. di questo volume.
Č importante quale gene della serie E determini la
colorazione di fondo. Per i galli il gene di base ha un’influenza
estremamente piccola: quando Db agisce sui fenotipi e+,
eb, ebc,
eWh, ey
si ottengono tutti soggetti dorati. Con il gene ER
si ottiene pur sempre un colore dorato, perň con remiganti che rimangono
nere, sia le primarie che le secondarie. Con E si hanno soggetti neri.
Nelle galline l’effetto esplicato da questo gene č
maggiore di quanto non si verifichi nel gallo:
§
con
eb le
galline ricevono una ripartizione dei colori di tipo columbia con strisce
sulla schiena e sul ventre, nonché un petto piů scuro
§
con
e+ presentano
una ripartizione columbia, perň il petto risulta color salmone
§
con
eWh
e con ey
si ottiene una colorazione del tutto simile alla
precedente.
Db, insieme a Co e a Ml, č
implicato nel disegno delle piume.
3. BASI GENETICHE DI ALCUNI FENOTIPI CON RESTRIZIONE COLUMBIA
Sebbene nelle restrizioni del
nero tipo columbia la quantitŕ di
eumelanina sia molto variabile, sono stati suggeriti tre gruppi:
columbia:
rosso
coda nera:
fulvo:

Fig. III. 6 - New Hampshire
Anche se attualmente č possibile identificare il genotipo
di base che giustifica le differenze tra i suddetti gruppi fenotipici nonché
le differenze in seno a ogni gruppo, esistono modificatori sconosciuti che determinano variazioni
cromatiche non direttamente legate agli alleli della serie E.
Nel 1966 Somes & Smyth si dedicarono
a una vasta serie di incroci utilizzando soggetti che differivano tra loro per
la quantitŕ di eumelanina posseduta da mantellina, ali e coda.
Ne dedussero che i fenotipi in studio differivano a causa di un controllo ereditario su base poligenica.
Le razze sottoposte a incrocio furono la Rhode Island red (la piů ricca di
nero), la New Hampshire (con nero in quantitŕ intermedia) e l’Orpington
fulva (priva di nero). Interessante il fatto che tutte e tre le razze avevano
in comune un genotipo di restrizione columbia: tutti i discendenti di F2 e delle successive generazioni mostravano infatti un
fenotipo columbia. Non era il caso del piumino, a carico del quale alcuni
fenotipi mostravano piů eumelanina di quanto si potesse osservare nelle linee
parentali.
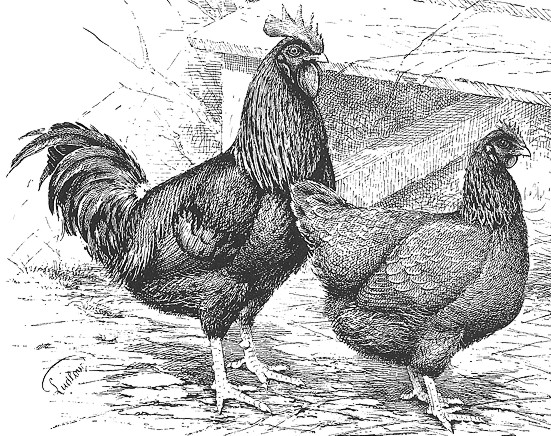
Fig. III. 7 - Sussex rossa. Da Edward Brown, 1906.
Ecco quali sono i genotipi ipotizzati per alcuni fenotipi
dotati di specifica restrizione columbia:
|
Columbia |
||||||
|
Bianco
columbia |
ebeb |
CoCo |
db+db+ |
mh+mh+ |
di+di+ |
SS |
|
Fulvo
columbia |
ebeb |
CoCo |
db+db+ |
mh+mh+ |
di+di+ |
s+s+ |
|
Rosso
columbia
[4]
|
ebeb |
CoCo |
db+db+ |
MhMh |
di+di+ |
s+s+ |
|
Giallo coda nera |
||||||
|
New Hampshire |
eWheWh |
CoCo |
db+db+ |
mh+mh+ |
di+di+ |
s+s+ |
|
Rosso coda nera |
||||||
|
Rhode Island Rossa |
eyey |
CoCo |
db+db+ |
MhMh |
di+di+ |
s+s+ |
|
Fulvo tipo Orpington |
||||||
|
Orpington fulva |
eWheWh |
CoCo |
db+db+ |
MhMh |
DiDi |
s+s+ |
Come giŕ accennato, al fulvo tipo Orpington appartiene la Cocincina fulva
![]() -
- ![]() -
- ![]() -
- ![]() .
.

Fig. III. 8 - Orpington fulva. Da Edward Brown, 1906.
|
Catalana del Prat Leonada |
|||||
|
Prat |
eWheWh |
Co/Co |
db+db+ |
mh+mh+ |
s+s+ |
Questa razza spagnola č caratterizzata da un piumino
arancio fulvo e da un piumaggio adulto ginger rosso. Il nome proviene da un’area
della Catalogna situata nei pressi di Barcellona: appena fuori Barcellona si
trova el Prat de Llobregat, con
65.000 abitanti.

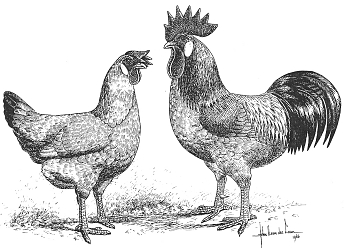
Fig. III. 9 - Catalana del Prat: a sinistra da Edward Brown (1906) - a destra dallo Standard olandese (1992).
Secondo Salvador Castelló, come riferisce Brown, si
tratterebbe di una razza originata dall’incrocio tra un ceppo indigeno e la
Cocincina. Ne scaturě un ibrido di grandi dimensioni, seguito poi da un
ritorno al tipo originario che corrisponde al ceppo Catalano. Si conservarono
solo le dimensioni ragguardevoli grazie a una selezione effettuata dagli
allevatori del Prat.
|
Vasca |
|||||
|
Vasca |
eWheWh |
CoCo |
db+db+ |
Mh/? |
s+s+ |
Gli adulti di questa razza sono piů scuri rispetto alla
Prat e somigliano alla New Hampshire. Le differenze nella tonalitŕ della
feomelanina pare siano dovute a un singolo gene che si comporta come dominante
nel dare una colorazione piů scura. Potrebbe trattarsi di alleli del locus
Co, oppure di qualche altro modificatore maggiore, forse il gene Mh.
|
Villafranquina |
|||||
|
Villafranquina |
ebeb |
co+co+ |
DbDb |
mh+/? |
s+s+ |
Si tratta di un’altra razza spagnola rossa coda nera
caratterizzata da un esteso disegno di tipo secondario presente sulla schiena
delle femmine, dovuto a Db.
|
Fulvo tipo Minorca |
||||||
|
Minorca fulva |
ey oppure ebc/? |
CoCo |
db+db+ |
MhMh |
DiDi |
s+s+ |
Anche questo fulvo, come quello
della Orpington, č una versione non eumelanotica della restrizione columbia.
I soggetti studiati erano eterozigoti per i due alleli della serie E.
Il
gallo di Ermanubi
il primo pollo color zafferano

Brabanter o Brabantea fulva testa nera
Il gallo, con la sua testa nera,
avrebbe fatto arricciare il naso a Ermanubi.
In greco
per designare il tuorlo dell'uovo - che č indubbiamente giallo, anche se con
tonalitŕ che variano fisiologicamente dal giallo-verde al croceo - si usa l'aggettivo ὠχρός (pallido, giallastro, come quando un soggetto č anemico), sostantivizzato nel neutro ὠχρόν: τὸ ὠχρόν
τοῦ ᾠοῦ - il giallo dell'uovo, Aristotele![]() Historia animalium 560a 21:
Historia animalium 560a 21:
"Il giallo e il bianco dell’uovo hanno natura opposta non solo per il colore ma anche per le loro proprietŕ. Il giallo infatti viene coagulato dal freddo, mentre il bianco non si coagula, anzi tende piuttosto a liquefarsi; sotto l’azione del fuoco il bianco coagula, il giallo no, anzi rimane molle a meno che non venga interamente bruciato, e viene condensato e disseccato piů dalla bollitura che dal fuoco vivo."
L'aggettivo
greco χλωρός in prima istanza significa
verde-giallo o verde-pallido, verdeggiante, verde, e la clorofilla č appunto
verde. Solo in seconda istanza significa giallo, e nelle vesti di sostantivo
neutro indica il giallo dell'uovo: ᾠοῦ τὸ
χλωρόν - il giallo
dell'uovo, come era
solito esprimersi il medico Zopiro citato da Oribasio![]() .
Il vocabolo greco fu usato nel 1810 da Sir Humphry Davy (Penzance 1778 -
Ginevra 1829) per battezzare un elemento gassoso a tutti noto – il cloro –
che č giallo-verde.
.
Il vocabolo greco fu usato nel 1810 da Sir Humphry Davy (Penzance 1778 -
Ginevra 1829) per battezzare un elemento gassoso a tutti noto – il cloro –
che č giallo-verde.
L'aggettivo
ὠχρός ha dato origine all'italiano ocra, termine generico usato per indicare
due minerali di ferro nella forma terrosa, l'ematite![]() e la limonite
e la limonite![]() ,
rispettivamente di colore rosso (ocra rossa) e di colore giallo (ocra gialla).
Finemente macinata, l'ocra viene utilizzata come pigmento per colori e
vernici. Mischiata con sostanze grasse, fu usata in etŕ preistorica - e lo č
tutt'oggi presso tribů primitive - per tingersi il corpo e il volto.
,
rispettivamente di colore rosso (ocra rossa) e di colore giallo (ocra gialla).
Finemente macinata, l'ocra viene utilizzata come pigmento per colori e
vernici. Mischiata con sostanze grasse, fu usata in etŕ preistorica - e lo č
tutt'oggi presso tribů primitive - per tingersi il corpo e il volto.
In greco
esiste un aggettivo per indicare in prima istanza il colore biondo o fulvo. Si
tratta di ξανθός, che viene per lo piů impiegato per i capelli, talora per la criniera
dei cavalli e per il miele, molto raramente per la criniera fulva del leone,
che solo dal poeta Oppiano di Apamea![]() fu etichettato come ξανθοκόμης, cioč dalla chioma bionda o, se vogliamo, dalla
fulva criniera.
fu etichettato come ξανθοκόμης, cioč dalla chioma bionda o, se vogliamo, dalla
fulva criniera.
Perň
grazie al Professor Antonio Garzya![]() č stato possibile appurare che il
geoponico Florentino
č stato possibile appurare che il
geoponico Florentino![]() (prima
metŕ del III sec. dC) si servě di ξανθός anche nel caso della livrea del pollo, e
precisamente - come vedremo tra poco - quando forně le caratteristiche
fenotipiche delle galline piů feconde: "generalmente
lo sono quelle di colore fulvo". Una caratteristica cromatica espressa da Florentino con αἱ
ξανθίζουσαι, nominativo femminile plurale del participio presente del verbo ξανθίζω, derivato ovviamente da ξανθός.
(prima
metŕ del III sec. dC) si servě di ξανθός anche nel caso della livrea del pollo, e
precisamente - come vedremo tra poco - quando forně le caratteristiche
fenotipiche delle galline piů feconde: "generalmente
lo sono quelle di colore fulvo". Una caratteristica cromatica espressa da Florentino con αἱ
ξανθίζουσαι, nominativo femminile plurale del participio presente del verbo ξανθίζω, derivato ovviamente da ξανθός.
Questo
aggettivo, che incontriamo nel termine xantofilla (il carotenoide![]() giallo presente nelle foglie, ma che fino all'autunno resta celato dalla verde
clorofilla), come tanti altri suoi colleghi di cromatologia antica possiede in
seconda istanza molteplici significati, spesso discordanti: giallo-verde,
verdastro, verde pallido, fino a significare biancastro, come dimostra
Teofrasto
giallo presente nelle foglie, ma che fino all'autunno resta celato dalla verde
clorofilla), come tanti altri suoi colleghi di cromatologia antica possiede in
seconda istanza molteplici significati, spesso discordanti: giallo-verde,
verdastro, verde pallido, fino a significare biancastro, come dimostra
Teofrasto![]() nel suo trattato sulle pietre.
nel suo trattato sulle pietre.

Xanthium
italicum – Lappolone – Compositae
foto
Elio Corti - 1972
Dioscoride![]() (IV,133) consigliava l'uso dello ξάνθιον per tingere di biondo i capelli: si tratta del
lappolone – Xanthium italicum – i cui frutti, conformati a oliva,
prima di seccare sono biondicci e giŕ armati di uncini, noti agli amanti di
passeggiate ecologiche, perché, essendo il lappolone una pianta zoofila, si
appiccicano tenacemente ai vestiti e al pelo del cane che ci accompagna,
assicurando cosě la disseminazione a distanza quando ripuliremo sia lui che
gli abiti.
(IV,133) consigliava l'uso dello ξάνθιον per tingere di biondo i capelli: si tratta del
lappolone – Xanthium italicum – i cui frutti, conformati a oliva,
prima di seccare sono biondicci e giŕ armati di uncini, noti agli amanti di
passeggiate ecologiche, perché, essendo il lappolone una pianta zoofila, si
appiccicano tenacemente ai vestiti e al pelo del cane che ci accompagna,
assicurando cosě la disseminazione a distanza quando ripuliremo sia lui che
gli abiti.

da Pierandrea Mattioli![]()
Commentarii in libros sex Pedacii Dioscoridis Anazarbei De Materia Medica
Venetiis, apud Valgrisium,
1554
Nel pollo
il gene dominante responsabile del biondo champagne![]() ,
un colore che richiama quello del lappolone, entra nella formula del piumaggio
fulvo, ma solo in dose singola, in quanto i soggetti omozigoti risultano poco
vitali. Per cui, anche se l'azione del gene Cb (champagne blond
- trovato per esempio nella Minorca fulva) sembra necessaria per diluire la
feomelanina in modo uniforme, č assai verosimile che non fosse presente nei
polli fulvi dell'antichitŕ, salvo lo fosse per caso o di straforo, in quanto
i soggetti omozigoti a un certo punto soccombevano. E a quei tempi non era
ancora certo giunto il momento di giocherellare come oggi con eterozigosi e
omozigosi: gli antichi dovevano produrre polli da combattimento e da tavola,
oppure ottime ovaiole, e basta.
,
un colore che richiama quello del lappolone, entra nella formula del piumaggio
fulvo, ma solo in dose singola, in quanto i soggetti omozigoti risultano poco
vitali. Per cui, anche se l'azione del gene Cb (champagne blond
- trovato per esempio nella Minorca fulva) sembra necessaria per diluire la
feomelanina in modo uniforme, č assai verosimile che non fosse presente nei
polli fulvi dell'antichitŕ, salvo lo fosse per caso o di straforo, in quanto
i soggetti omozigoti a un certo punto soccombevano. E a quei tempi non era
ancora certo giunto il momento di giocherellare come oggi con eterozigosi e
omozigosi: gli antichi dovevano produrre polli da combattimento e da tavola,
oppure ottime ovaiole, e basta.
Ξάνθος, che per puri motivi di accento sembrerebbe non aver nulla a che fare
con ξανθός, credo invece abbia molto da condividere con questo aggettivo. Ξάνθος č il nome di diversi fiumi cosě chiamati
nell'antichitŕ, il piů famoso dei quali fu lo Scamandro, detto anche Xanto,
che col Simoenta - nati ambedue dal Monte Ida oggi Kazdag - lambiva Troia. E
potremmo fare un'illazione: magari uno Ξάνθος aveva spesso le acque gialle, come il cinese Fiume
Giallo (Hwang Ho) che trasporta enormi quantitŕ di detriti, tra cui il
giallastro löss![]() ,
tanto da tingere di giallo, insieme ad altri fiumi minori, anche il mare in
cui sfocia: il Mare Giallo (Hwang Hoi), un settore di ben 417.000 km2 dell'Oceano Pacifico a nord del Mar Cinese
Orientale.
,
tanto da tingere di giallo, insieme ad altri fiumi minori, anche il mare in
cui sfocia: il Mare Giallo (Hwang Hoi), un settore di ben 417.000 km2 dell'Oceano Pacifico a nord del Mar Cinese
Orientale.
Quel che
č certo č che ξανθός non venne mai usato per esprimere una livrea biondo
champagne. Il veto era puramente biologico e di origine genetica. Č assai
probabile che Florentino abbia usato il verbo ξανθίζω con lo stesso significato espresso da Plutarco![]() attraverso il sostantivo maschile κροκίας, cioč, color zafferano e non biondo champagne.
attraverso il sostantivo maschile κροκίας, cioč, color zafferano e non biondo champagne.
Per
indicare una livrea fulva del pollo - piů precisamente una livrea color
zafferano![]() - solo in Plutarco
(ca. 46 - ca. 125) troviamo il sostantivo maschile κροκίας. Egli č l'unico autore a riferire con questo termine la colorazione del
piumaggio di un pollo che ancora nel II secolo dC veniva immolato a Ermanubi
- solo in Plutarco
(ca. 46 - ca. 125) troviamo il sostantivo maschile κροκίας. Egli č l'unico autore a riferire con questo termine la colorazione del
piumaggio di un pollo che ancora nel II secolo dC veniva immolato a Ermanubi![]() .
.
E siccome
Ermanubi fu partorito poco dopo che Alessandro Magno![]() prese possesso dell'Egitto (332 aC), possiamo dedurre che questa colorazione
del piumaggio - e tutti i relativi geni implicati - fosse giŕ presente nel
mondo egiziano qualche secolo prima che Plutarco ne parlasse.
prese possesso dell'Egitto (332 aC), possiamo dedurre che questa colorazione
del piumaggio - e tutti i relativi geni implicati - fosse giŕ presente nel
mondo egiziano qualche secolo prima che Plutarco ne parlasse.
Per inciso
ricordiamo che pare essere stato il faraone Taharqa![]() - della XXV dinastia, morto nel 664 aC - a favorire in Egitto l'avicoltura che
si trasformň in fonte di reddito nazionale. A questo periodo risalgono i
famosi megaincubatoi
- della XXV dinastia, morto nel 664 aC - a favorire in Egitto l'avicoltura che
si trasformň in fonte di reddito nazionale. A questo periodo risalgono i
famosi megaincubatoi![]() che ai tempi di Réaumur
che ai tempi di Réaumur![]() (1683-1757), piů precisamente intorno al 1750,
sfornavano circa 93 milioni di polli l'anno. Ma J. B. Coltherd (The domestic fowl in
ancient Egypt, Ibis n° 108, 1966) č piů dell’avviso che i primi veri tentativi di un allevamento
intensivo del pollo nella Valle del Nilo vadano datati ai tempi dei Saiti
(quando Sais, situata sul ramo di Rosetta del Delta del Nilo, fu capitale del
regno dal 663 al 525 aC durante la XXVI dinastia, detta appunto saita o
saitica) o, piů probabilmente, che vadano datati ai tempi dei Tolomei
(1683-1757), piů precisamente intorno al 1750,
sfornavano circa 93 milioni di polli l'anno. Ma J. B. Coltherd (The domestic fowl in
ancient Egypt, Ibis n° 108, 1966) č piů dell’avviso che i primi veri tentativi di un allevamento
intensivo del pollo nella Valle del Nilo vadano datati ai tempi dei Saiti
(quando Sais, situata sul ramo di Rosetta del Delta del Nilo, fu capitale del
regno dal 663 al 525 aC durante la XXVI dinastia, detta appunto saita o
saitica) o, piů probabilmente, che vadano datati ai tempi dei Tolomei![]() .
.
Aristotele
non menzionň mai polli dalla livrea fulva nel mondo greco, né lo fecero per
il mondo romano Catone![]() ,
Varrone
,
Varrone![]() ,
Plinio
,
Plinio![]() e Columella
e Columella![]() ;
solo piů tardi li menzionň Palladio
;
solo piů tardi li menzionň Palladio![]() (IV sec. dC) in Opus agriculturae
libro I, XXVII (De gallinis) parlando telegraficamente delle galline: “Sint
praecipue nigrae, aut flavi coloris, albae vitentur.”, cioč, siano
soprattutto nere, o di colore giallo (giallo vivo, giallo oro, aranciato,
biondo), si evitino quelle bianche.
(IV sec. dC) in Opus agriculturae
libro I, XXVII (De gallinis) parlando telegraficamente delle galline: “Sint
praecipue nigrae, aut flavi coloris, albae vitentur.”, cioč, siano
soprattutto nere, o di colore giallo (giallo vivo, giallo oro, aranciato,
biondo), si evitino quelle bianche.
Appena prima di Palladio - ma per l'area greca, e circa un secolo e mezzo dopo Plutarco – fu Florentino a parlare di polli fulvi - αἱ ξανθίζουσαι, come segnalato dal Professor Antonio Garzya - e ce ne dŕ notizia Gessner a pagina 424 della sua ornitologia (1555):
Gallinas educaturus eligat foecundissimas: quas nimirum ex usu rerum et experientia dignoscet: im<m>o vero ex pluribus aliis indiciis. In universum enim quae colore flavescunt, et sortiuntur digitos impares, quaeque magna possident capita (τὰς ὄψεις μεγάλας, oculos magnos, Cornarius) cristamque erigunt: nec non nigriores et corpulentiores. Eae omnes gallinae facile mares ferent: multo erunt praestantiores ad partum, ova maxima {a}edent: ac breviter, generosos excludent pullos, Florentinus.
Chi dovrŕ allevare delle galline scelga le piů
feconde: senza dubbio sarŕ in grado di riconoscerle in base alla pratica e
all’esperienza: anzi, in base a numerosi altri indizi. Infatti generalmente
lo sono quelle di colore fulvo, e che ricevono in sorte le dita
dispari, e quelle che hanno la testa grande (tŕs ópseis megálas, gli
occhi grandi, in base alla traduzione di Janus Cornarius![]() )
e che tengono la cresta dritta: nonché quelle che sono di colore piů scuro e
che sono piů corpulente. Tutte queste galline sopporteranno facilmente i
maschi: saranno di gran lunga superiori riguardo alla deposizione, faranno
delle uova molto grandi: e in breve volgere di tempo daranno alla luce dei
pulcini di buona qualitŕ, Florentino.
)
e che tengono la cresta dritta: nonché quelle che sono di colore piů scuro e
che sono piů corpulente. Tutte queste galline sopporteranno facilmente i
maschi: saranno di gran lunga superiori riguardo alla deposizione, faranno
delle uova molto grandi: e in breve volgere di tempo daranno alla luce dei
pulcini di buona qualitŕ, Florentino.
Non č questo il momento di entrare in polemica con
Gessner e con Cornarius, secondo i quali Florentino avrebbe affermato che le
galline di razza si distinguono per avere la testa grossa (Gessner) o gli
occhi grossi (Cornarius). Per i dettagli circa l'esatta interpretazione del
testo di Florentino si veda quanto specificato nel lessico![]() .
.
Per il gallo di Ermanubi era in ballo un piumaggio croceo, color croco, cioč color zafferano - tra il giallo e il rosso - o, come puntualizza Filippo Capponi in Per un lessico tecnico pliniano (1991), un giallo che tende al rosso. Plinio usň l'aggettivo croceus parlando della starna (come Capponi consiglia di emendare in X,134: Est et alia nomine eodem, a coturnicibus magnitudine tantum differens, croceo {unctu} <tinctu> cibis gratissima.), ma giammai usň croceus a proposito di una livrea del pollo.
Quindi si
puň presumere che per il gallo di Ermanubi si trattasse di una colorazione giŕ
presente nei polli egiziani, ma ignota fino al IV secolo dC in quelli romani,
come possiamo desumere da Palladio. Per Florentino possiamo supporre che egli
conoscesse la pentadattilia![]() del pollo in quanto dal Nordeuropa questa mutazione aveva raggiunto Roma e poi
la Grecia, ma forse i suoi polli fulvi avevano prima raggiunto il mondo greco
e poi quello romano provenendo dall'Egitto.
del pollo in quanto dal Nordeuropa questa mutazione aveva raggiunto Roma e poi
la Grecia, ma forse i suoi polli fulvi avevano prima raggiunto il mondo greco
e poi quello romano provenendo dall'Egitto.
Κροκίας deriva da κρόκος che
significa zafferano (Crocus sativus), e se il sostantivo κρόκος viene usato da Alessandro di Tralles![]() (527-565) col significato di tuorlo d'uovo, per il lessicografo Esichio di
Alessandria
(527-565) col significato di tuorlo d'uovo, per il lessicografo Esichio di
Alessandria![]() (V sec dC) κρόκος corrisponde
al gallo con il collo color zafferano, quindi un gallo rosso-nero tipo Gallus
gallus oppure un gallo con mantellina – e con la sola mantellina - color
arancio per diluizione del rosso grazie al gene dell'argento.
(V sec dC) κρόκος corrisponde
al gallo con il collo color zafferano, quindi un gallo rosso-nero tipo Gallus
gallus oppure un gallo con mantellina – e con la sola mantellina - color
arancio per diluizione del rosso grazie al gene dell'argento.
Quindi il κρόκος di Esichio non identifica un gallo interamente color zafferano come accade invece per κροκίας di Plutarco. Giŕ alcuni secoli prima del lessicografo alessandrino il nostro Columella in De re rustica VIII,2,9 parlava come lui di mantellina fulva, ma solo di mantellina e non di tutta quanta la livrea: iubae deinde variae vel ex auro flavae, per colla cervicesque in umeros diffusae - piume del collo variegate o dorate tendenti al giallo, che si diffondono per tutto il collo fino alle spalle.
In italiano non abbiamo l'abitudine di dire piumaggio croceo (giallo tendente al rosso, come puntualizzato da Capponi) ma parliamo di piumaggio fulvo, aggettivo derivato dal latino fulvus che significa colore biondo o giallo rossiccio tipico della criniera del leone. Quindi, se non vogliamo esaurire i nostri poveri neuroni, possiamo trovare un concordato: dire croceo o dire fulvo o dire leonino č la stessa cosa, come dimostra il linguaggio catalano nel definire la colorazione della razza del Prat: Catalana del Prat leonada. E i Francesi dicono fauve.
Tanto per portare due esempi sull'uso di fulvus per designare il colore del leone, ecco due famosi poeti latini:
Virgilio![]() Georgiche IV,408: fulva cervice leaena – la leonessa dal collo fulvo.
Georgiche IV,408: fulva cervice leaena – la leonessa dal collo fulvo.
Lucrezio![]() De rerum natura 5,901: corpora fulva leonum – i corpi fulvi dei
leoni.
De rerum natura 5,901: corpora fulva leonum – i corpi fulvi dei
leoni.
Gli anglofoni - e anche gli Olandesi - per identificare il piumaggio fulvo usano l'aggettivo buff – color camoscio - che deriva dal sostantivo buff corrispondente alla pelle del bisonte americano, detto buffalo (Bison bison), il povero bovide imparentato con buoi e bufali che immortalň Buffalo Bill – alias William Frederick Cody (contea di Scott, Iowa, 1846 - Denver, Colorado, 1917) – il quale si impegnň a rifornire gli operai della ferrovia del Pacifico di carne di bisonte: sosteneva di aver ucciso piů di 4.000 bisonti - in americano buffalo - in meno di diciotto mesi.
In tedesco la colorazione fulva č detta molto semplicemente gelb, cioč gialla.
A questo punto spero vi stiate domandando come mi sia stato possibile incappare nel testo di Plutarco relativo al pollo color zafferano immolato a Ermanubi. State pur certi che non mi sono mai preso né mai mi prenderň la briga di leggere per intero Iside e Osiride, una delle opere che compongono la collezione dei Moralia di Plutarco.
Il merito č tutto di Gessner, e per riflesso di Aldrovandi.
A pagina 402 di Historia animalium III (1555) il grande medico zurighese comincia a titillare la nostra curiositŕ adducendo le citazioni di Esichio e Plutarco:
|
Κρόκος, τὸ
κροτόν, (mendum est forte:) et
gallinacei qui collum habent eiusmodi, (croceis vel aureis iubis
scilicet ornatum,) κρόκη,
Hesych. et Varinus. Gallus κροκίας
Hermanubidi immolabatur, Plutarchus. |
Krókos
– zafferano, tň krotón (forse questo č un errore): sono
pure dei galli che hanno il collo di questo colore (cioč il cui
collo č ornato da mantelline color zafferano oppure color oro), krókë
– mantello, Esichio e Guarino. A Ermanubi veniva immolato un gallo
krokías – color
zafferano, Plutarco. |
A pagina 407 Gessner č piů dettagliato:
|
Anubis apud
Aegyptios vocatur (vocabulo Graecae originis) ὁ
ἀναφαίνων
τὰ οὐράνια
καὶ τῶν ἄνω <φερομένων>
λόγος:
hoc est ratio superiorum et coelestia declarans. Et idem interdum
Hermanubis, quod nomen rebus inferioribus convenit, ut illud
superioribus. Sacrificant autem utrique gallum, illi album, quod
coelestia syncera [sincera] et lucida existiment: huic κροκίαν,
(hoc est pennis et iubis croceis praeditum, Gyraldus etiam croceum
transtulit,) inferiora omnia mixta et varia esse rati, Plutarchus in
lib. de Iside et Osiride. Pyrrhus rex cum splene laborantibus
mederetur, albo gallo sacrum peragebat, Lilius Gr. [408] Gyraldus. |
Anubi |
Aldrovandi fa le stesse citazioni di Gessner, rispettivamente a pagina 188 e 256 del II volume di Ornithologia (1600).
Quindi: tutti i geni implicati nel piumaggio fulvo – con le opportune limitazioni per il gene Cb - erano giŕ presenti in polli egiziani durante il periodo ellenistico, quando sbocciň Ermanubi.

Buff
Cochins
Il gallo non avrebbe fatto arricciare il naso a Ermanubi.
L'immagine č dovuta al talento artistico di J. W. Ludlow
ed č contenuta in The Illustrated Book of Poultry (1890) di Lewis
Wright (1838-1905).
4.
altri TIPI CON RESTRIZIONE COLUMBIA molto
eumelanizzati
Quaglia e Lakenveld
Parecchie
varietŕ columbia si manifestano in una versione fortemente eumelanizzata. Una
versione melanotica della Prat
eWh/eWh_Co/Co
č stata trovata omozigote per il gene Ml.

Fig. III. 10 - Brabançonne o Pollo del Brabante
Lo stesso č stato suggerito anche per la colorazione quaglia delle Nane Belghe e della
Brabançonne. Secondo Jeffrey nella colorazione quaglia il colore di base
sarebbe dovuto a e+
allo stato omozigote, al quale č sufficiente l’associazione con Co e con un
fattore eumelanizzante sconosciuto per ottenere la caratteristica
distribuzione dei pigmenti.
Sono tuttavia necessarie ulteriori ricerche per chiarire
la formula genetica della colorazione quaglia. Per ora ci si deve accontentare
di quanto afferma Carefoot: uno stato omozigote per Co, Ml e pg+, associato a e+
oppure a eb, rappresenta un corredo
genetico essenziale, anche se incompleto, di questa diffusa colorazione ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() .
.
Il fenotipo della Lakenveld, definibile come una
restrizione columbia fortemente eumelanizzata, dotata del gene S al posto del
gene s+, dovrebbe possedere un genotipo
eb/eb
Co/Co cui si aggiungono geni eumelanizzanti non identificati, capaci di
segregare per il gene Ml ![]() -
- ![]() . In assenza del gene Co, ma coi geni
eumelanizzanti presenti, la Lakenveld dŕ luogo al fenotipo nero. La Vorwerk
č la versione feomelanica del mantello della Lakenveld.
. In assenza del gene Co, ma coi geni
eumelanizzanti presenti, la Lakenveld dŕ luogo al fenotipo nero. La Vorwerk
č la versione feomelanica del mantello della Lakenveld.
Digressione storica sulla Lakenveld
Viene fatta menzione della
Lakenveld in un documento di viaggio datato 1727. Il nome deriverebbe dal
villaggio olandese di Lakenveld, situato tra Lexmond e Meerkerk, a sud di
Utrecht, oggi denominato Lakerveld. Un’altra ipotesi vuole che la razza
debba il nome alla disposizione dei pigmenti del piumaggio: il nero del collo
e della coda viene avvolto dal bianco del tronco come se fosse un lenzuolo,
detto laken in olandese. Una
colorazione identica č presente nei bovini Lakenveld. Si suppone che questi
polli juventini siano scaturiti da
soggetti con barratura autosomica presenti da secoli in tutta l’Europa Nordoccidentale: Campine, Brakel, Pollo della Frisia, Gabbiano della Frisia
Orientale. Ed č proprio per la grande diffusione di tali razze barrate nell’Europa
Nordoccidentale che i Tedeschi reclamano la paternitŕ della Lakenveld, che
intorno al 1850 era diffusamente allevata in Westfalia e nelle campagne di
Hannover.
Non certo per creare confusione, ma solo per dimostrare
come le notizie si perdano talora nella notte dei tempi, aggiungiamo che in
base alle ricerche di Brown la Lakenveld sarebbe di origine orientale: questo
studioso le assegna come patria Gerusalemme.

Fig. III. 11 - Polli e bovini Lakenveld
5. ALTRI GENI DI RESTRIZIONE tipo COLUMBIA
Senza dubbio esistono altri geni
che tendono a restringere l’eumelanina del petto e/o del mantello. Desta
meraviglia il fatto che alcuni di essi sembrano essere ipostatici rispetto ai
geni associati del nero esteso. Cote (1976)
su nove maschi di razze tra loro diverse, nonché omo o eterozigoti per il
gene E, ha trovato 8 soggetti che possedevano modificatori tipo columbia particolarmente attivi a livello del petto.
Questi casi mostrarono una considerevole variazione di
penetranza e di espressivitŕ. Nessuno dei soggetti era portatore di Db e solo
2
possedevano
il gene Co. Uno di questi ultimi era un Plymouth Rock barrato da esposizione,
e questo fatto č molto interessante alla luce delle osservazioni che il gene Co
intensifica l’espressione del barrato legato al sesso
dotato di uno sfondo e+/eb
.
|
sommario |
top |
avanti |
|
|
|