Vol. 3° - IV.
I GENI RESPONSABILI
DEL DISEGNO
secondario del piumaggio
|
|
Un tempo si riteneva che la responsabilitŕ dei diversi
disegni secondari del piumaggio ricadesse su un singolo gene. Seguendo questo postulato
non era perň possibile spiegare le numerose varianti. Si ipotizzň allora l’intervento
di geni modificatori capaci di agire
sull’unico gene responsabile del disegno.
Recentemente č stato dimostrato che sono in gioco piů
geni e che la loro azione combinata č in grado di determinare un disegno
specifico. Abbiamo giŕ incontrato un trio di geni coinvolti nel disegno
secondario delle piume - Co-Db-Ml - dotati anche di un effetto sulla distribuzione delle
melanine, cioč sul disegno primario. Rimangono ancora da analizzare mo e Pg.
Per ogni tipo di disegno, eccetto il punteggiato, risulta
necessaria la presenza del gene Pg. Se Pg č l’unico gene in causa,
determina il pluriorlato.
Tutti i geni del disegno secondario - eccetto mo - si comportano come
dominanti, e sono tutti autosomici. Esiste una circostanza molto particolare: Co-Db-Ml-Pg si trovano tutti quanti sullo
stesso cromosoma, sul cromosoma 1. Sono quindi associati.
Ecco cosa ha proposto Clive
Carefoot ![]() :
:
|
Interazioni
geniche responsabili |
||||||
|
Fenotipo |
serie E | Pg/pg+ | Co/co+ | Db/db+ | Ml/ml+ | Mo+/mo |
| Punteggiato | e+ | pg+ | co+ | db+ | ml+ | Mo+ |
| Pluriorlato | eb | Pg | ||||
| Barrato autosomico | ||||||
| tipo Amburgo | eb | Pg | Db | |||
| tipo Fayoumi | ER | Pg | Co | Db | ||
|
tipo Buttercup
[1]
|
ebc | Pg | Db | |||
| Orlo singolo | eb | Pg | Co | Ml | ||
| Orlo doppio | eb, eWh, ey | Pg | Ml | |||
| Pagliettato | E o ER? | Pg | Db | Ml | ||
| Pomellato | E | ? | ? | ? | mo | |
| Millefiori | eb o eWh? | ? | Co | ? | mo | |
Incrociando soggetti dotati di
geni appartenenti a questa categoria, bisogna tener presente che nei
discendenti il rapporto numerico tra i differenti tipi di disegno sarŕ
diverso da quanto ci si aspetterebbe dalle leggi mendeliane. Bisogna inoltre
ricordare che il colore dorato selvatico non possiede geni del disegno. La sua
formula č: pg+
db+
ml+
co+
Mo+.
L’azione dei geni che analizzeremo si esplica su tutti
gli alleli della serie E, in quanto sono essi a determinare la distribuzione e l’arrangiamento delle melanine nel
contesto del mantello e della piuma. Molti geni agiscono sia su
sfondo oro che su sfondo argento, a loro volta condizionati dalla presenza o
dall’assenza di feomelanina nella parte non eumelanizzata della piuma.
Tutti
quanti sono dotati di una certa attivitŕ barrante,
che si manifesta lungo l’asse maggiore della piuma al momento della
formazione delle barbe. Il meccanismo genetico che accende e spegne la
melanizzazione varia da un disegno all’altro: un continuo accendi e spegni
si verifica per le barre bianche e nere della Plymouth Rock, si ha una sola
accensione e un solo spegnimento dell’interruttore eumelanizzante nel caso
dell’orlo singolo della Wyandotte.
Esistono delle particolaritŕ da segnalare. Il barrato
autosomico - o barrato trasversale, o pluriorlato parallelo, detto pencilling
[2]
o pencilled
nell’Amburgo e barrato nella
Campine - a differenza del barrato legato al sesso non possiede alcun effetto
sulle strutture dell’iporachide. La paillette
dell’Amburgo deve essere rotondeggiante in Inghilterra, mentre negli USA si
pretende che sia a forma di lettera V. Nel piumaggio giovanile le copritrici alari presentano
un disegno quasi simile in tutte le combinazioni geniche - cioč una barratura
irregolare ![]() - e in tutte le svariate combinazioni non si nota
alcuna influenza particolare sull’iporachide.
- e in tutte le svariate combinazioni non si nota
alcuna influenza particolare sull’iporachide.
La maggior parte dei disegni secondari č su base
autosomica. Le revisioni e gli aggiornamenti in questo campo della genetica
sono stati innumerevoli: per fare un esempio, č stato dimostrato che l’orlo
semplice della Wyandotte si puň spiegare attraverso il contributo di 3 geni,
mutanti di 3 loci differenti. Un altro esempio del livello raggiunto nell’analisi
genica č quello del pomellato, dovuto al solo mutante mo che puň dar
luogo a ben 5 fenotipi diversi. La maggior parte dei recenti progressi deriva
dall’identificazione sia degli alleli della serie E che delle mutazioni
implicate nella restrizione del nero, nonché dalla comprensione di alcune
interazioni fra tutti questi geni.
Il contributo alla genetica non proviene solo da studiosi
che l’hanno scelta come prima passione. Infatti la lotta ingaggiata in
campo genetico dall’inglese Carefoot, espositore e giudice di polli
ornamentali nonché uomo d’affari laureato in matematica, lo dimostra appieno. Egli
ha sovvertito tutta la genetica dei disegni. Per fare un ultimo esempio, egli
ha proposto che il gene Pg č il gene del pagliettato.
I geni piů significativi implicati in certi disegni sono
stati dimostrati con sufficiente certezza. Bisogna tuttavia tenere ben
presente che č necessaria l’identificazione
di altri geni modificatori per giungere a fenotipi di qualitŕ elevata.
Č stato dimostrato che il gene Pg appartiene al III
gruppo di linkage sul cromosoma 1 e che č associato al gene Db e al gene Ml,
approssimativamente e rispettivamente distanziati 20 e 10 unitŕ crossover,
con interposto il locus Ml.
Per
ulteriori particolari si veda III-1.1. - Melanotico
1. Punteggiato e pepato
|
pg+_e+ |
Il
Gallo Rosso della giungla presenta uno spiccato dicromatismo sessuale. In alcune zone sessualmente
dimorfiche il
piumaggio dei maschi contiene aree
nere associate a feomelanina. Le femmine presentano una fine punteggiatura
marrone scuro e marrone chiaro, non perň a livello del petto salmone, che non
č punteggiato ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() .
.
Il piumino di questi pulcini č caratterizzato da evidenti
strisce al capo, al dorso e ai fianchi, pigmentate in marrone scuro e disposte
su sfondo bianco giallastro. In assenza di modificatori maggiori, come Co
oppure Db, la colorazione dell’adulto č quella appena descritta. Il piumino
a strisce del pulcino e il petto salmone della femmina suggeriscono l’azione
di un solo gene. Tuttavia, la punteggiatura puň riscontrarsi anche in altri
genotipi, come per esempio eb
che differisce dal tipo selvatico per il colore del piumino e per l’assenza
del salmone al petto della femmina adulta. Nel 1985 Carefoot
ha suggerito che una singola mutazione, cioč Pg, č in grado di alterare il
processo di punteggiatura, tanto da consentire un certo numero di
riarrangiamenti dell’eumelanina nel contesto della piuma, riarrangiamenti
concentrici che derivano da modificazioni geneticamente determinate.
Sino a questo momento non č dato sapere se lo sviluppo o
l’assenza di punteggiatura possano essere attribuiti a un singolo gene. In
base alle conoscenze attuali la presenza
di punteggiatura puň essere designata col simbolo pg+.
La pepatura causata da e+
consiste in puntini piccolissimi che si possono osservare sulla schiena delle
femmine dorate, a condizione che non siano presenti geni del disegno, ma
solamente e+
- pg+
- co+
- db+
- ml+
- Mo+.
Allo
scopo di ottenere dei punti molto piccoli nelle femmine
si dovrebbe disporre di un maschio caratterizzato dal minor numero possibile
di lanceolate ampiamente nere sia al collo che alla sella e dotato di
punteggiatura nel triangolo dell’ala. Tale maschio va incrociato con una
femmina che si avvicini il piů possibile allo standard. Se si usa un maschio
che presenta delle chiazze marroni o delle orlature al petto, si otterranno
delle femmine con disegni che sembrano piccole scintille o orlature, forse
dovute a uno stato eterozigote per uno dei geni implicati nel disegno e che
ora analizzeremo.
Fig. IV. 1 - Mappa del linkage del cromosoma 1
2. PLURIORLATO e BARRATO AUTOSOMICO
Questa
terminologia viene spesso confusa dagli allevatori, che talora fanno
riferimento ai due fenotipi come se anche il secondo fosse un disegno
pluriorlato. Poveretti, essi non hanno tutti i torti, visto che c’č chi ha
avuto la pensata di definire il barrato autosomico anche pluriorlato parallelo, forse con lo scopo di distinguerlo dal
barrato ovale anch'esso autosomico detto anche barrato
diagonale.
2.1. Pluriorlato
|
Pg
- patterning - |
Autosomico
incompletamente dominante
Gruppo di associazione III - cromosoma 1
Il
disegno pluriorlato della femmina consiste in una serie di ellissi
concentriche e tra loro separate; il margine della piuma ha un bordo dello
stesso colore di quello delimitato dalle ellissi
![]() -
- ![]() -
- ![]() -
- ![]() .
I maschi hanno un piumaggio eminentemente di tipo selvatico senza appariscenti
influssi da parte di Pg.
.
I maschi hanno un piumaggio eminentemente di tipo selvatico senza appariscenti
influssi da parte di Pg.
Come gene del disegno
secondario, nel pluriorlato č presente solo Pg, che il piů delle volte
esplica i suoi effetti su uno sfondo con genotipo e+ o eb. Nel caso che il genotipo sia
diverso, gli effetti di Pg sono simili ma meno netti, in quanto i risultati
migliori si ottengono su uno sfondo legato a e+ oppure a eb piuttosto che su uno
sfondo determinato da ey.

Fig. IV. 2 - Wyandotte pluriorlata
2.2. Barrato autosomico
|
Pg
+ Db |
Pg
- autosomico incompletamente dominante
Db - autosomico incompletamente dominante,
influenzato dal sesso in E
I due geni appartengono al gruppo di associazione III
- cromosoma 1
Il
barrato autosomico č anche detto pluriorlato parallelo.
Č ormai definitiva la convinzione secondo cui questo disegno non č affatto
dovuto a un singolo gene Ab, simbolo
usato in passato come acronimo di autosomal
barring.
Un soggetto pluriorlato si trasforma in barrato quando
viene aggiunto il gene Db. La piuma presenta cosě delle strisce disposte
trasversalmente rispetto al rachide, ed essendo questa barratura dovuta a geni
autosomici, essa viene trasmessa in modo identico sia alla discendenza
maschile che femminile. Per cui non avremo un mantello differenziato in base
al sesso come invece accade per il barrato legato al sesso. Altra differenza č la
mancanza di barratura all’iporachide ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() .
.
A seconda che il colore di sfondo sia dovuto a ER, e+,
eb oppure al frumento, l’effetto barrato č piů o meno
intenso. Oltretutto si puň ottenere una modificazione della barratura:
infatti, se ai geni Pg e Db fa da sfondo ebc
oppure ey, la barratura assume la forma di una striscia
composta da due chiazze ovali giustapposte e si ottiene lo spruzzato, o barrato
ovale, o barrato diagonale, o listato.

Fig. IV. 3 - Amburgo spruzzata oro
Il disegno spruzzato - o listato - caratteristico di Amburgo spruzzata, Brakel, Bresse, Hergnies, Landaise, č dotato di penetranza e di espressione variabili. Proprio a causa della sua penetranza variabile, il disegno sia dei maschi che delle femmine di Bresse grigia variano da un soggetto all’altro. Lungi dall’essere un’anomalia contro cui l’allevatore deve lottare, questa eterogeneitŕ del piumaggio č normale. Pertanto la comparsa di barre sulle ali č difficile da ottenere e la loro presenza sta sě a testimoniare il talento dell’allevatore, ma soprattutto il capriccio della genetica. A questo proposito bisogna sottolineare che la rappresentazione di un piumaggio perfetto in una Bresse grigia č piů spesso un disegno anziché una fotografia.
Quest’esigenza
di una perfezione del barrato, che non č ottenibile in modo regolare,
rappresenta una delle cause, forse la principale, del declino della Bresse
grigia. Bisogna dunque che l’allevatore di questa varietŕ sappia
comportarsi con umiltŕ di fronte a un disegno talora sconcertante e che
consideri lo standard un fine verso il quale deve tendere la sua paziente
selezione, certamente non come un imperativo da raggiungere a ogni costo. Lo standard
meriterebbe una revisione in tal senso.
2.3. Considerazioni pratiche sul pluriorlato e sul barrato autosomico
Č interessante notare che il piumaggio
giovanile
- sia
del barrato autosomico che del pluriorlato
- č caratterizzato da un identico disegno barrato.
Ambedue i disegni consistono in un riarrangiamento dell’eumelanina che,
nella forma classica, si esprime su uno sfondo feomelanico. Sia il pluriorlato
che il barrato autosomico sono dei genotipi caratterizzati da un dimorfismo
sessuale: si esprimono nella femmina di qualsiasi etŕ, mentre nel maschio questi disegni si esprimono solo nel piumaggio giovanile,
a meno che il maschio adulto non sia caratterizzato da un piumaggio gallina.
Ambedue i fenotipi hanno un piumaggio giovanile del tutto simile,
caratterizzato da un’esile barratura trasversale. Lo sviluppo del
pluriorlato dal barrato ha inizio nel piumaggio giovanile tardivo, quando
presenta delle barrature solo all’apice delle piume, barrature che col
passare del tempo si fanno via via piů numerose e sempre piů curve sino a
raggiungere il pluriorlato dell’adulto.
Per le femmine giŕ Kimball aveva proposto che un gene Pg,
incompletamente dominante, č in grado di riorganizzare la punteggiatura
dando origine a un disegno pluriorlato. Carefoot ha aggiunto un ruolo piů
ampio a tale gene: le varietŕ da lui testate si erano dimostrate dotate del
gene eb, e anche se egli non č
a conoscenza di varietŕ e+ pluriorlate, si aspetterebbe che le femmine adulte
e+Pg/e+Pg
abbiano piume pluriorlate su tutto il mantello, eccetto che al petto salmone.

Fig. IV. 4
- Brakel spruzzata oro
Il gallo ha un disegno del mantello come quello della gallina
Nel 1985 Carefoot giunse alla conclusione
che il barrato autosomico differisce dal pluriorlato per l’aggiunta - a un
genotipo non punteggiato Pg/Pg - di un gene di restrizione tipo columbia, e
precisamente del gene Db allo stato omozigote. Il gene Db sarebbe l’induttore
del barrato ed esistono relazioni di linkage tra Db e Pg: i
due geni sono separati da circa 20
unitŕ
crossover. In passato si disse che questa stessa distanza separava Db e Ab,
per cui quest’ultimo gene non č altro che il gene Pg.
Č stata dimostrata la presenza di Db nel barrato
autosomico della Buttercup, caratterizzato da una barratura particolare,
essendo di tipo ovale. Il gene Db č stato riscontrato anche in un gallo
Amburgo spruzzato oro, che č un barrato autosomico. Polli Fayoumi studiati da
Smyth avevano una colorazione di base determinata da ER/ER, mentre nei soggetti di Carefoot la Buttercup possedeva
il suo gene ebc, e l’Amburgo possedeva un
genotipo eb/eb.

Fig. IV. 5 - Il barrato ovale della Buttercup
Se si vuol scendere in particolaritŕ che mettono in
risalto quanto sia talora complessa la genetica del barrato autosomico, si
puň precisare che uno stato omozigote per Db e per eb
produce una barratura in cui il nero č due volte piů esteso del rosso.
Questo sta a significare una regola generale: a
seconda del gene della serie E
presente, nonché a seconda della sua etero/omozigosi, possono mutare le
proporzioni tra il nero e il rosso. Infatti sembra probabile che
il barrato migliore si ottenga nelle femmine con genotipo eb_ebc.
Al contrario, per ottenere dei maschi con una bella coda nera, č auspicabile
un genotipo ebc_ebc,
che nella femmina equivalente comporta una notevole riduzione del nero sul
mantello, ed č verosimile che Pg non sia presente, oppure č meglio che sia
assente per ottenere i migliori risultati ai fini di un’esposizione, che
cozzano sempre contro la purezza genetica. Ma l’allevatore talora puň,
anzi, deve eludere la purezza genetica, in quanto attraverso il pollo deve
esprimere il suo gusto estetico derivato dall’esperienza.
Che l’esperienza sia di valido supporto alla scienza č
dimostrato dal fatto che č estremamente difficile allevare femmine Amburgo
spruzzata oro dotate di un buon bilanciamento fra nero e rosso. Se una femmina
č ben barrata al petto, tende ad avere la schiena troppo scura e a presentare
troppo nero sulla mantellina. Se poi la barratura č regolare, il petto č
troppo chiaro. Non č nota la base genetica della differenza tra le barre
parallele dell’Amburgo e delle macchie
ovali della Buttercup, ma si presume vengano implicati alcuni geni modificatori.
Gli allevatori cercano di arrivare al risultato desiderato miscelando questi
effetti. In altre parole, non fanno altro che giostrare coi geni modificatori,
che pur essendo dotati di un’azione minore non sono assolutamente d’importanza
secondaria.
Le difficoltŕ nel classificare i fenotipi che segregano
da incroci in cui sono coinvolti il punteggiato, il pluriorlato e il barrato
autosomico, hanno indotto Carefoot a usare dei ceppi da esposizione
relativamente omogenei, in modo da semplificare la classificazione dei
discendenti. Nella generazione F2
derivante da un maschio perniciato puro omozigote eb/eb_pg+/pg+ con femmine Plymouth Rock pluriorlate ma molto variabili
nella qualitŕ del pluriorlato, in molti casi le figlie erano di difficile
classificazione. Risultati variabili sono stati ottenuti in F2 anche dall’incrocio tra un Sebright argento, che ha un
orlo semplice, e femmine Amburgo gold
pencilled che sono barrate autosomiche. Le variazioni del pluriorlato si
notano soprattutto a livello delle ali e alla base della coda.
3. ORLATO
Attualmente si ritiene che non
esista un gene specifico per l’orlatura, denominato in passato Lg
che sta per lacing. Questo gene č sostituito oggi da altri 2 o 3 geni:
§ orlo
doppio: intervengono i geni
Pg e Ml
§
orlo
singolo: intervengono
i geni Pg, Ml e Co
Ambedue
gli effetti dell’orlatura debbono esplicarsi su uno sfondo e+, eb,
eWh oppure ey.
3.1. Orlo doppio
|
Pg
+ Ml |
Pg
- autosomico incompletamente dominante
Ml - autosomico incompletamente dominante
I due geni appartengono al gruppo di associazione III
- cromosoma 1
Se
si prende un soggetto pluriorlato puro e+/e+_Pg/Pg e lo incrociamo con un
soggetto melanotico e+/e+_Ml/Ml, si ottengono dei discendenti con orlo
doppio, per cui in questo disegno sono implicati i geni Pg e Ml. Le penne
hanno un orlo scuro esterno e uno interno la cui larghezza č influenzata da
un altro gene per ora ignoto. Lo spazio tra i due orli ovviamente non č
eumelanotico. In certe piume ciň che rimane di un terzo orlo si addossa al
rachide ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() .
.
La combinazione di questi geni quasi non si nota nei
maschi, soprattutto se i geni agiscono su uno sfondo e+ oppure eb: infatti i galli presentano un piumaggio di tipo selvatico.
Con un colore di base e+,
talora il maschio presenta orli sul petto. Le femmine presentano invece un
doppio orlo.
Dal momento che Pg e Ml sono dominanti, anche se in modo incompleto, il fenotipo dei soggetti non puri č identico a
quello degli omozigoti.
Si č visto che anche splendide femmine Cornish posseggono un orlo singolo alla gola e un triplo orlo sopra al cuscino e nella parte bassa del petto. Spesso nella zona del cuscino esiste un bordo dello stesso colore dello sfondo, posto all’estrema periferia della piuma, al di lŕ dell’orlo nero esterno. Inoltre, sempre in questa zona, talora la percentuale di nero č talmente scarsa da generare l’effetto di un disegno pluriorlato. Non foss’altro che per semplici motivi visivi, appare chiara la relazione tra il pluriorlato e il biorlato. Ambedue i disegni presentano anelli concentrici e paralleli al bordo della piuma, con una sola differenza: il pluriorlato deve avere il colore di sfondo all’estrema periferia, dopo l’ultimo orlo nero, mentre la periferia di una piuma con doppio orlo inizia subito con un orlo nero intensamente pigmentato. Un’altra differenza consiste nella mantellina, che si presenta disegnata nella femmina pluriorlata, mentre nella Cornish č spesso praticamente nera, come accade anche nella Barneveld.
Allora viene spontaneo chiedersi se la differenza
principale tra i due disegni non consista semplicemente nella dotazione di un
gene che incrementa in modo drammatico la quantitŕ di nero. La risposta č
sě. Si tratta del gene Ml.

Fig. IV. 6 - Barneveld
In passato č stato possibile dimostrare che la Cornish č dotata di uno dei due geni
frumento come alternativa al gene posseduto dal tipo selvatico. Questa
constatazione fu confermata dalla comparsa di pulcini paglierini nella seconda
generazione derivata da un incrocio tra Wyandotte perniciata e Cornish. Tale
colore chiaro comparve a dispetto del fatto che i geni intensificatori del
nero presenti nella Cornish sarebbero di per sé in grado di causare delle
strisce nere a carico del piumino di questi pulcini.

Fig. IV. 7 - Barneveld coi suoi pulcini
La Barneveld č l’esempio migliore di doppia orlatura con
sfondo eb, e il suo maschio non č cosě
intensamente melanotico come il Cornish dotato di sfondo eWh. Verosimilmente nel Cornish esistono altri geni in
grado di intensificare il nero a un punto tale da fargli assumere un aspetto
veramente tetro. Per molti anni la Cornish č stata inincrociata allo scopo di
ottenere il colore che lo standard pretende. Cosě facendo si compromette il
colore e il disegno, in quanto si va inevitabilmente incontro a un
deterioramento della colorazione per voler ricavare ambedue i sessi da uno
stesso gruppo di riproduttori. Accade pertanto di vedere soggetti dalla forma
splendida ma dal colore deludente.
3.2. Orlo singolo
|
Pg
+ Co + Ml |
Pg
- autosomico incompletamente dominante
Co - autosomico incompletamente dominante
Ml - autosomico incompletamente dominante
I tre geni appartengono al gruppo di associazione III
- cromosoma 1
L’orlo
singolo č un derivato dell’orlo doppio per aggiunta del gene Co ai geni Pg
e Ml. Per effetto del columbia, che rimuove l’eumelanina dal centro della
piuma, le due bande del doppio orlo si trasformano in una striscia singola
piů larga e periferica. Anche se questi 3 geni sono dominanti, l’orlo
risulta migliore se essi sono allo stato omozigote.
L’orlo
peggiore, caratteristico dei soggetti eterozigoti,
deriva dalla presenza di un
solo gene Co.
La presenza singola dei due altri geni non determina alterazioni dell’orlo.
L’abolizione del gene Co dal genotipo permette al fenotipo doppio orlo di riesprimersi.
L’orlo singolo č piů evidente nelle femmine, in quanto sono dotate di
orlatura su tutte le penne, mentre nel maschio l’orlo č spesso presente
solo al petto e al ventre.
Il disegno a orlo singolo tende a concentrare l’eumelanina
al bordo della piuma, che ha un colore differente rispetto al centro
![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() .
Tale bordo č spesso nero, ma puň essere bianco, blu oppure splash. Questo
disegno esiste nell’Andalusa, nella Sebright, in certe varietŕ di Padovana
e di Wyandotte, piů raramente nella Cocincina nana o gigante. A perfezionare
la Cocincina nana argento orlo nero, giŕ descritta negli USA, si stanno
dedicando i fratelli Maurizio e Luciano
Tona di Casatenovo (LC).
.
Tale bordo č spesso nero, ma puň essere bianco, blu oppure splash. Questo
disegno esiste nell’Andalusa, nella Sebright, in certe varietŕ di Padovana
e di Wyandotte, piů raramente nella Cocincina nana o gigante. A perfezionare
la Cocincina nana argento orlo nero, giŕ descritta negli USA, si stanno
dedicando i fratelli Maurizio e Luciano
Tona di Casatenovo (LC).

Fig. IV. 8 - Sebright argento
Qualora l’orlo singolo fosse nero, la piuma si presenta
contornata da un anello periferico di eumelanina. L’area interna, non nera,
č bianca oppure oro, a seconda dell’allele presente nel locus S. Come
abbiamo appena detto, questo disegno č posseduto da ambedue i sessi, ma č
diverso nelle regioni del piumaggio sessualmente dimorfiche, in quanto nel
gallo solo il petto e il ventre sono orlati in modo tipico. La presenza di un
piumaggio di tipo femminile Hf/Hf, come nella Sebright, produce un piumaggio
completamente orlato in ambedue i sessi.
Non solo a Carefoot sarŕ capitato spesso di osservare in
Wyandotte nane argento orlo nero in periodo
depositivo, meritevoli peraltro di un primo premio, che la piuma
ricresce con un secondo
orlo interno ![]() -
- ![]() .
Questa chiazza sovente č mal distinguibile, ma talora č ben evidente. Un
effetto del tutto simile puň verificarsi anche nella varietŕ oro orlo nero,
in cui č talora presente anche una diluizione del colore dorato centrale.
Quasi tutte le femmine delle due varietŕ, ultimata la muta, diventano non
idonee a una competizione a causa del nero che si č aggiunto al centro delle
piume, quello che spesso gli allevatori definiscono muschiositŕ.
.
Questa chiazza sovente č mal distinguibile, ma talora č ben evidente. Un
effetto del tutto simile puň verificarsi anche nella varietŕ oro orlo nero,
in cui č talora presente anche una diluizione del colore dorato centrale.
Quasi tutte le femmine delle due varietŕ, ultimata la muta, diventano non
idonee a una competizione a causa del nero che si č aggiunto al centro delle
piume, quello che spesso gli allevatori definiscono muschiositŕ.

Fig. IV. 9 - Padovana oro orlo blu
Carefoot ha voluto verificare se in quest’alterazione
del mantello possano intervenire fattori ormonali. Di proposito egli ha messo
a covare su uova finte una femmina oro orlo nero e le permise di rimanervi per
nove settimane, in capo alle quali la chioccia abbandonň spontaneamente
il nido. La
gallina andň incontro a muta totale e sviluppň un piumaggio talmente ben
orlato da vincere il primo premio nell’Esposizione Nazionale Inglese del 1978.
Due anni piů tardi lo stesso esperimento fu tentato con la sorella e il
risultato fu identico.
Sempre Carefoot ha osservato che talora la Cornish, dotata
di orlo doppio, presenta un orlo singolo alla gola. Probabilmente, laddove l’entitŕ
della pigmentazione č debole, il doppio orlo perde il suo nucleo centrale,
mentre al dorso, dove la pigmentazione č abitualmente piů intensa, l’orlatura
si presenta doppia. Partendo dal presupposto che l’orlo singolo č dotato
dei geni Ml, Co e Pg, č possibile trasformare un orlo doppio in singolo
aggiungendo alla sua formula due geni Co? Ed č possibile l’inverso,
togliendo i due geni columbia?
Per saperlo č necessario incrociare un rappresentante di
ciascun disegno tra loro e quindi incrociare i discendenti tra loro. L’esperimento
ha potuto rispondere di sě alla seconda domanda, e quindi per illazione č
valido anche l’inverso.
Il fatto di sapere che i due tipi di orlatura differiscono
per intervento di un solo gene, il columbia, deve indurre gli allevatori
perfezionisti ad astenersi dall’incrociare soggetti con orlo semplice e
doppio. Le Wyandotte orlo blu dalla mantellina uniforme e i soggetti dotati di
muschiositŕ rappresentano classici esempi delle difficoltŕ che dovremo
affrontare per recuperare un collo orlato una volta che esso sia andato perso
per incroci eseguiti alla carlona.
La femmina di Sebright ha la coda dotata di orlatura,
mentre la Wyandotte orlata possiede una coda nera. Pertanto deve essere
presente un fattore capace di restringere il nero nella coda della Sebright.
Mentre l’ereditarietŕ dell’orlatura della Sebright non č stata ancora
analizzata in modo soddisfacente, si puň tuttavia supporre che la coda
orlata, nonché lo sfondo piů chiaro nella varietŕ oro, siano dovuti a
qualche fattore del fulvo non ancora identificato.
3.3. L’orlatura della coda nella Sebright secondo Lewis Wright
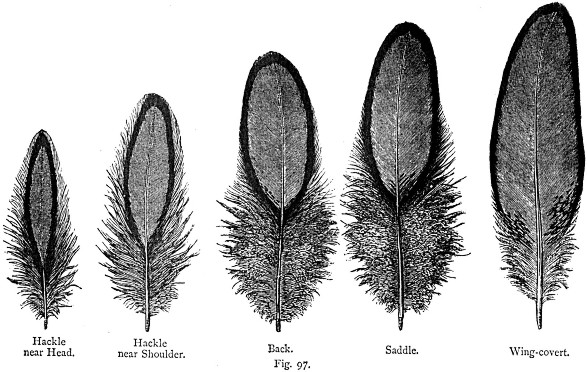
Orlatura
senza difetti delle piume di Sebright secondo Lewis Wright
eccettuate
quelle della coda
The illustrated book of Poultry (1890)
Nella Sebright le piume piů predisposte a errori di orlatura sono quelle della coda, dove l’orlo si presenta frequentemente sottile in corrispondenza dei bordi laterali. Una frequente constatazione di Lewis Wright, frutto di piů di vent’anni di lavoro, era quella della corrispondenza tra orlo peggiore e orecchione di un bianco perfetto [3] . Non esiste una ragione evidente di questo parallelismo, puntualizza Wright; tuttavia č prudente selezionare per l’orlatura quei soggetti che presentano un orecchione magari scadente, sacrificando le caratteristiche dell’orecchione a quelle di una bella orlatura.
Ma bisogna tenere presente un altro
suggerimento al fine di ottenere un orlato shocking:
i soggetti migliori si ottengono da genitori dei quali uno presenta un orlo
ampio e l’altro un orlo tenue appena sufficiente. Dobbiamo infine ricordarci
di un altro comandamento dettato a fine ottocento: mentre č concesso
incrociare occasionalmente Sebright oro e argento per migliorare la varietŕ
oro, i soggetti argento che nascono da questo incrocio, anche se idonei a
essere esposti alle mostre, non debbono assolutamente essere accoppiati con
qualsivoglia ceppo argento puro.
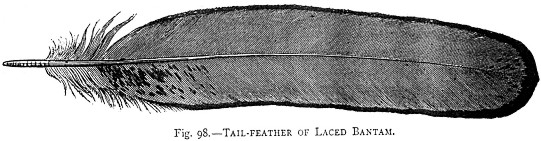
Orlatura
difettosa delle piume della coda di Sebright secondo Lewis Wright
The illustrated book of Poultry (1890)
Wright, avendo scoperto che la
scarsa fertilitŕ della Sebright dovuta alla necessitŕ di uno stretto inbreeding
puň essere ridotta incrociando i soggetti con la Nana di Giava, ha tuttavia
potuto osservare che il nero della seconda impartisce una tonalitŕ
eccessivamente rossa alla feomelanina della Sebright oro.
3.4. Curriculum vitae dell’orlatura semplice e doppia
Alla luce delle recenti
acquisizioni, la genetica dell’orlato risulta abbastanza complessa. Nel 1972
Smyth
aveva riscontrato nella Wyandotte argento orlo nero la presenza di uno stato
omozigote per 4 geni: eb,
Co, Ml, Lg. I loci Ml e Lg
avevano un linkage di circa 10 unitŕ crossover. Successivamente Lg si dimostrň essere null’altro che Pg.
Precedenti osservazioni avevano dimostrato che l’orlo
semplice si evidenziava solo in piume nelle quali l’eumelanina era stata
ristretta dal gene Co, e fu verificato da Smyth che il gene Co era necessario
per l’espressione dell’orlo singolo. Il gene Lg da solo causava un’orlatura incompleta che veniva relegata alla
punta della piuma. Il gene Ml era richiesto per completare l’orlo, in quanto
i due geni funzionavano in modo dose-dipendente.
Nel 1986 Carefoot confermň quest’ultima
osservazione di Smyth, ma chiarě che il gene Lg altro non era se non quello che egli aveva giŕ denominato Pg.
Perciň affermň che Co č l’istigatore dell’orlatura semplice attraverso
l’interazione con Pg, e l’anno seguente riuscě a dimostrare che i loci Pg
e Ml hanno la stessa relazione di linkage di quella stabilita per Lg
e Ml da parte di Smyth.
Anche la genetica del doppio orlo č stata sovvertita
dalle ricerche di Carefoot: non si tratta dell’azione di Lg in associazione con Pg su di uno sfondo e+/e+,
dove il gene Pg era responsabile dell’orlo interno. La rimozione di Co da un
genotipo eb/eb
Co/Co Ml/Ml Pg/Pg caratterizzato da un orlo singolo, dava luogo a femmine con
doppio orlo e a maschi di tipo selvatico.
Il disegno a doppio orlo della Barneveld e quello a orlo
unico della Wyandotte hanno come sfondo un genotipo eb/eb, ma lo stesso disegno puň manifestarsi anche su uno
sfondo frumento, come il frumento recessivo ey
della Cornish scura, o fagianata, che in un’altra linea dimostrň essere
invece eWh.
Sempre da Carefoot, nel 1988,
č stato dimostrato che l’orlo singolo dell’Andalusa blu č dovuto a un
genotipo Co-Ml-Pg, cui perň fa da sfondo l’allele E.
3.5. Origine del nome Wyandottes
Come c’era da aspettarsi, l’etimologia
č incerta. Deriverebbe da quello di una tribů di Indiani del Nordamerica, un
tempo numerosa, nota prima col nome di Wandots, Wiandots, Wayandots,
Weyondottes, Wiondots, Wiyandottes e finalmente Wyandottes. La storia dice che
la penisola compresa tra i laghi Huron, Erie e Ontario - dove si trova
Toronto, tanto per intenderci - era occupata da due popolazioni distinte che
parlavano dialetti della lingua Irochese. Gli Uroni, o Wyandottes, abitavano
le foreste della riva orientale del lago al quale hanno dato il loro nome, il
Lago Huron. I Wyandottes erano in parte agricoltori. Guerreggiarono a lungo
coi bellicosi Irochesi, finché nel 1648 si ebbe lo scontro finale con
successivo abbandono da parte dei Wyandottes della terra natale. Dopo varie
peripezie, questo popolo trovň pace e fondň un nuovo villaggio nell’area
di Sandusky, Ohio.
Č stato scritto che l’omonima razza di polli avrebbe
preso il nome dalla regione in cui fu inizialmente allevata, appartenente alla
tribů dei Wyandottes, ma Houdlette precisň che la scelta del nome attribuito
ai polli ha ben altra origine, in quanto egli era particolarmente affezionato
all’imbarcazione guardacoste usata da suo padre, battezzata Wyandottes. (Da:
Standard Bred of Wyandottes -
Reliable Poultry Journal Pub.Co. - Quincy - Illinois - 1910)

Fig. IV. 10 - Wyandotte argento orlo nero
4. Pagliettato
Partiamo
dall’esperienza per spiegare meglio le nuove teorie circa questo disegno, le
quali hanno come presupposto il fatto che il barrato autosomico č dovuto all’azione
congiunta di Pg e Db, e non all’azione di un singolo gene Ab.
Se un barrato autosomico omozigote viene incrociato con un soggetto nero
dotato di Ml, quindi nero profondo, i discendenti saranno dei pagliettati, in
quanto il disegno pagliettato proviene dalla combinazione di Pg, Db, Ml. Tutti
e 3 i geni si trovano sullo stesso cromosoma e sono associati, o linked.
Pagliettato
|
Pg
+ Db + Ml |
Pg
- autosomico incompletamente dominante
Db - autosomico incompletamente dominante,
influenzato dal sesso in E
Ml - autosomico incompletamente dominante
I tre geni appartengono al gruppo di associazione III
- cromosoma 1
Visto
che i 3 geni sono tutti dominanti, dall’incrocio suddetto possono scaturire
soggetti eterozigoti per il disegno pagliettato. Se si pratica un incrocio tra
soggetti pagliettati omozigoti, il trio di geni pare comportarsi come un gene
singolo.
Fino a non molti anni fa si pensava che il pagliettato
dipendesse da un solo gene siglato Sp
- spangled
- e per comoditŕ
potremmo continuare a usare questa terminologia diventata ormai obsoleta.
Sp
agisce bene sia su sfondo argentato che dorato. Caratterizza Ciuffata di
Appenzell o Appenzeller spitzhauben ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() , Barbuta di Turingia
pagliettata, Amburgo pagliettata, siano esse argentate che dorate.
, Barbuta di Turingia
pagliettata, Amburgo pagliettata, siano esse argentate che dorate.
Questo disegno consiste in un lustrino di eumelanina a
forma di lettera V aperta all’esterno secondo lo standard USA ![]() -
- ![]() , ma rotondeggiante secondo gli Europei
, ma rotondeggiante secondo gli Europei ![]() ,
il quale č localizzato nella parte distale della piuma. La restante porzione,
come nel caso dell’orlato, puň essere bianca oppure colorata dalla
feomelanina. Le varietŕ oro e argento dell’Amburgo differiscono non solo
per il colore dell'area non pagliettata, ma anche per il fatto che le timoniere, le falciformi
e le copritrici della coda sono nere nella varietŕ oro, mentre sono bianche
con la punta fornita di lustrino nella varietŕ argento. La ragione di questa
differenza non č nota. Le femmine delle due varietŕ hanno un lustrino piů
ampio del maschio, forse dovuto a un’eumelanogenesi accentuata dagli
estrogeni piuttosto che per qualche gene legato al sesso (Taylor, 1932).
,
il quale č localizzato nella parte distale della piuma. La restante porzione,
come nel caso dell’orlato, puň essere bianca oppure colorata dalla
feomelanina. Le varietŕ oro e argento dell’Amburgo differiscono non solo
per il colore dell'area non pagliettata, ma anche per il fatto che le timoniere, le falciformi
e le copritrici della coda sono nere nella varietŕ oro, mentre sono bianche
con la punta fornita di lustrino nella varietŕ argento. La ragione di questa
differenza non č nota. Le femmine delle due varietŕ hanno un lustrino piů
ampio del maschio, forse dovuto a un’eumelanogenesi accentuata dagli
estrogeni piuttosto che per qualche gene legato al sesso (Taylor, 1932).

Fig.
IV. 11
- Ciuffata di Appenzell o Appenzeller
spitzhauben
pagliettata argento
Il
piumino dei pulcini pagliettati č caratterizzato da strisce dorsali
irregolari, o imperfette. Il piumino
della testa nella varietŕ argento č essenzialmente chiaro, mentre nella
varietŕ dorata mostra una striscia di estensione variabile. Sul piumino di
questi pulcini il pagliettato non si esprime e si rende manifesto solo a circa
4 settimane d’etŕ ![]() -
- ![]() .
.

Fig. IV. 12 - Amburgo pagliettata argento
Come si č detto, un’interpretazione meno recente
attribuě questo disegno a un singolo gene Sp.
Si tratta di studi del passato abbastanza confusi e solo ultimamente Carefoot
ha fatto luce nel tunnel intricato dello spangling:
si tratterebbe di un effetto combinato di Db e Ml, che conducono alla
concentrazione di eumelanina in un lustrino posto all’apice della piuma. Si
tratterebbe di un conflitto tra il restrittore del nero Db e l’intensificatore
del nero Ml. In occasione dell’orlo semplice abbiamo detto che Ml si combina
con Co per determinare la formazione di un singolo orlo marginale, e anche in
questo caso si tratterebbe di un conflitto tra i geni Co e Ml, che cosě
riescono a determinare l’orlatura. Da ciň si č dedotto che il
pagliettato č dovuto a un’omozigosi dei geni co+
Db Ml Pg.
Il pagliettato si esprime sia su sfondo E che su sfondo eb
[4]
,
anche se nel secondo caso il lustrino risulta molto piccolo (Carefoot, 1985).
Perň, come fa notare Smyth, nessuno ha finora isolato e studiato il gene
della colorazione di base sul quale agiscono i geni del disegno nel dare il
pagliettato, per vedere se il gene del locus E, un gene simil-E, non sia per caso
un betulloide, capace di essere
piegato ai voleri espressivi dei geni del disegno. Altrimenti, dove si va a
pescare la feomelanina affinché venga separata dall’eumelanina? E su chi
agirebbe il gene dell’argento se non disponesse del suo substrato, l’oro?
Personalmente non sono d’accordo sulla presenza dell’allele
E allo stato omozigote. Altrimenti dobbiamo rivedere e riarrangiare tutte le
nostre idee sull’espressione degli alleli della colorazione di base, pur
essendo consci che il nero integrale si ottiene con l’aggiunta di Ml. Il
gene Ml a braccetto con eb
dŕ i soggetti neri a zampe gialle. Questa colorazione dei tarsi sarebbe un’utopia
se l’allele fosse E. Perň, da come abbiamo martellato finora, se vogliamo
dei soggetti neri senza una favilla di rosso dobbiamo ricorrere all’allele
E.
|
Si č giŕ detto che, per spiegare la differenza tra la barratura ovalare della Buttercup e quella dell’Amburgo spruzzata, si ipotizza l’intervento di geni modificatori. Lo stesso concetto č applicabile per spiegare il disegno pagliettato a mezzaluna caratteristico della Redcap. Da notare che il disegno di questo piumaggio č difficile da maneggiare, in quanto non sempre si ottengono i risultati desiderati a carico della mezzaluna: il punto critico sta nel miscelare adeguatamente i modificatori minori. |
|
Un’altra mutazione che puň avere un’azione modificatrice sul pagliettato
č il gene Mh, dotato di un effetto pagliettato-simile sul petto nero dei maschi di tipo selvatico.
Questo gene verosimilmente č presente nell’Amburgo pagliettata oro, in cui
determina anche la caratteristica tonalitŕ mogano della feomelanina.
L’Amburgo pagliettata argento non č l’esatta
controparte della varietŕ dorata. Nella pagliettata argento la coda deve
avere le paillettes e i maschi
debbono portare sulla mantellina delle macchie a forma di stiletto. Finora non
č dato sapere se sono necessari altri fattori di restrizione oppure se il gene S
č anche dotato di un’azione di restrizione sul nero
con riduzione della quantitŕ di nero depositata sulla piuma, per cui le aree
non nere si fanno piů estese. Non bisognerebbe meravigliarsi qualora a carico
del gene dell’argento venisse dimostrato quest’effetto di restrizione, in
quanto si č potuto verificare che tutti i geni di restrizione del nero
determinano un’alterazione dell’intensitŕ e della tonalitŕ della
feomelanina contemporaneamente presente. Forse č questa la vera ragione per
cui č piů facile ottenere una bella coda in una Sebright argento rispetto
alla varietŕ oro.
Le differenze nel piumino sono ben evidenti se il gene Db č presente oppure
assente. Allo scopo di definire il piumino del pulcino pagliettato sono stati
studiati soggetti con cresta a pisello e soggetti a collo nudo. Per i geni in
causa si č potuta dimostrare la seguente relazione di linkage: P-32-Db-10-Ml-10-Pg-26-Na,
e questo corrisponde con le osservazioni del passato, anche se il problema era
stato esposto in altro modo, adducendo l’esistenza di un gene del marmorizzato,
dato che talora i pulcini presentavano un piumino appunto marmorizzato, dovuto a Db. Questo aspetto del piumino presuppone un
genotipo eb.
Circa la striscia al capo, presente in modo caratteristico
nei pulcini dell'Amburgo pagliettata oro, pare sia dovuta a un’interazione di s+,
e ciň non deve destare meraviglia, in quanto anche le code degli adulti oro e
argento sono differenti.
Abbiamo giŕ detto che, se un barrato autosomico puro
viene incrociato con un soggetto nero melanotico, i discendenti saranno dei
pagliettati, in quanto il pagliettato proviene dalla combinazione di Pg, Db,
Ml. Tutti e tre i geni si trovano sullo stesso cromosoma e sono associati, o linked.
Siccome questi tre geni son tutti dominanti, č possibile che un soggetto
pagliettato non sia obbligatoriamente omozigote per tutti e tre.
Se un pagliettato puro viene incrociato con un soggetto
non pagliettato, la prole sarŕ tutta pagliettata. Successivamente, se si
pratica un reincrocio, si ottengono tanto dei pagliettati quanto dei non
pagliettati. I risultati dipendono in gran parte dal genotipo del genitore non
pagliettato. Se il genitore non pagliettato č dotato di Pg, esiste una
probabilitŕ maggiore che i figli presentino il pagliettato rispetto a quando
il genitore non possiede Pg. In un soggetto con piume disegnate č in ogni
caso presente il gene Pg, necessario anche per il pagliettato.
In un barrato autosomico sono
giŕ presenti Pg e Db. Per cui in un incrocio tra pagliettato e barrato
autosomico si ottengono piů pagliettati rispetto a un incrocio tra
pagliettato e pluriorlato.
Vediamo di comprendere meglio
questi concetti servendoci del quadrato di Punnett.
| Maschio pagliettato | Femmina con barratura autosomica | ||
|
eb eb_Pg Pg_Ml Ml_Db Db |
eb eb_Pg Pg_ml+ ml+_Db Db |
||
| eb =
perniciato Pg = gene del disegno Db = dark brown ml+ = non estensione melanica, allele di Ml di estensione melanica in
F1 avremo 100% di soggetti pagliettato impuro analizziamo
F2 derivante da |
|||
| 75%
pagliettato 25% barrato autosomico |
gameti femminili |
||
| eb Pg Ml Db | eb Pg ml+ Db | ||
|
gameti maschili |
eb Pg Ml Db | eb
Pg Ml Db eb Pg Ml Db pagliettato |
eb
Pg Ml Db eb Pg ml+ Db pagliettato |
| eb Pg ml+ Db | eb
Pg ml+ Db eb Pg Ml Db pagliettato |
eb
Pg ml+ Db eb Pg ml+ Db barrato |
|
| Maschio pagliettato | Femmina pluriorlata | ||||
|
eb eb_Pg Pg_Ml Ml_Db Db |
eb eb_Pg Pg_ml+ ml+_db+ db+ |
||||
| eb =
perniciato Pg = gene del disegno Db = dark brown e db+ suo allele ml+ = non estensione melanica, allele di Ml di estensione melanica F1: pagliettato impuro con formula eb eb_Pg Pg_Ml ml+_Db db+ F2:
pagliettato impuro x pagliettato impuro |
|||||
| 56,25%
pagliettato 18,75% doppio orlo 18,75% barrato autosomico 6,25% pluriorlato |
gameti femminili |
||||
| Ml Db | Ml db+ | ml+ Db | ml+ db+ | ||
|
gameti maschili |
Ml Db | Ml Db Ml Db pagliettato |
Ml Db Ml db+ pagliettato |
Ml Db ml+ Db pagliettato |
Ml Db ml+ db+ pagliettato |
| Ml db+ | Ml
db+ Ml Db pagliettato |
Ml
db+ Ml db+ due orli |
Ml
db+ ml+ Db pagliettato |
Ml
db+ ml+ db+ due orli |
|
| ml+ Db | ml+ Db Ml Db pagliettato |
ml+ Db Ml db+ pagliettato |
ml+
Db ml+ Db barrato |
ml+ Db ml+ db+ barrato |
|
| ml+ db+ | ml+
db+ Ml Db pagliettato |
ml+
db+ Ml db+ due orli |
ml+
db+ ml+ Db barrato |
ml+
db+ ml+ db+ pluriorlato |
|
Anche se Pg, Db e Ml sono
associati, in realtŕ, con gli incroci esemplificati, non si ottiene una
segregazione secondo le leggi mendeliane.
Se considerassimo questi tre geni come un solo gene Sp,
incrociando tra loro dei pagliettati non omozigoti dovremmo aspettarci una
segregazione secondo i principi di Mendel: 25% pagliettato puro, 50%
pagliettato impuro e 25% non pagliettato. Ma questo risultato non si verifica
e ora vedremo il perché.
I soggetti non puri possono
essere designati con la formula: Pg_pg+/Db_db+/Ml_ml+. Durante la meiosi i geni Pg e
pg+,
Db e db+,
Ml e ml+
possono scambiarsi tra loro. Questo fenomeno dello scambio di geni fra due
cromosomi omologhi va sotto il nome di crossingover, che giŕ conosciamo. A
causa del crossingover si riduce la percentuale di associazione tra Db-Ml-Pg,
per cui si otterranno meno discendenti pagliettati di quanto ci si
aspetterebbe da una segregazione mendeliana, valida qualora si adottasse il
concetto di un gene unico. La probabilitŕ di una dissociazione č sempre piů
piccola quanto piů i geni si trovano tra loro vicini.
4.1.a. Amburgo - Fagiano dello Yorkshire o Semilunati del Lancashire - Redcap
I polli che in Inghilterra
andavano sotto il nome di Amburgo avevano perlomeno due distinte origini. L’Amburgo
spruzzata fu importata dall’Olanda ed era chiamata
Dutch Everyday Layer o anche Everlasting
Layer. Al contrario l’Amburgo sia pagliettata che nera debbono essere
considerate come razze nate in Inghilterra molto tempo fa, la prima nota da
sempre come Lancashire Mooneys e Yorkshire
Pheasant, mentre la varietŕ nera andava sotto il nome di Black
Pheasant Fowl. Si deve verosimilmente al Reverendo Dixon il loro
raggruppamento sotto il nome onnicomprensivo di Amburgo, senza rispettare
perň i dati storici, geografici e somatici, in quanto si tratta di polli
simili ma che presentano alcune non trascurabili differenze.

Fig. IV. 13 - Redcap
Brown, che ha filtrato i dati di Wright e di Beldon,
giunge alla conclusione condivisa da Watchorn che la Redcap proviene dall’incrocio
tra l’Amburgo pagliettata oro e il Combattente inglese antico. Un tempo la
Redcap - detta da Pascal![]() Gallina a cuffia
rossa
- era molto comune nello Yorkshire, specialmente nella zona collinare
che si trova al sud della contea, nonché nel Derbyshire, cosicché ne
esistono almeno due varietŕ, ma secondo altri allevatori le varietŕ sono tre,
distinte in base alle dimensioni della caratteristica cresta. Chi possiede la
cresta di volume maggiore č la varietŕ dello Yorkshire, mentre la Derbyshire ha una
cresta di medie dimensioni. A queste due si affianca la varietŕ con cresta
piccola che non ha un nome specifico.
Gallina a cuffia
rossa
- era molto comune nello Yorkshire, specialmente nella zona collinare
che si trova al sud della contea, nonché nel Derbyshire, cosicché ne
esistono almeno due varietŕ, ma secondo altri allevatori le varietŕ sono tre,
distinte in base alle dimensioni della caratteristica cresta. Chi possiede la
cresta di volume maggiore č la varietŕ dello Yorkshire, mentre la Derbyshire ha una
cresta di medie dimensioni. A queste due si affianca la varietŕ con cresta
piccola che non ha un nome specifico.
|
sommario |
top |
avanti |
|
|
|
[1] Il tipo Buttercup del barrato autosomico puň anche essere denominato spruzzato, o barrato ovale, o barrato diagonale, o listato.
[2] Attenzione a non confondere pencilling e pencilled del barrato, dagli identici termini usati per definire il pluriorlato. Credo che gli statunitensi preferiscano una elle sola, mentre gli inglesi ne usano due. Certo che l’armamentario di vocaboli usato da un fancier č piů da crazy che da persona normale! Vien quasi da rimpiangere i tempi in cui ogni allevatore se ne stava a casa sua e non doveva fare il saltimbanco per adeguarsi alle esigenze linguistiche delle mostre estere che visitiamo sempre piů spesso. Concedetemi un’ultima cosa: il barrato legato al sesso č forse verticale? No, č solo non diagonale, mentre č diagonale la barratura ovale della Siciliana.
[3]
The
illustrated book of Poultry (1890) - Oggi lo standard vuole un orecchione rosso, pur concedendo un
colore turchese, che, come nella Silky ![]() , č un colore strutturale dovuto al
bianco, a sua volta determinato dalle guanine.
, č un colore strutturale dovuto al
bianco, a sua volta determinato dalle guanine.
[4] I geni di sfondo sono stati dedotti dalle formule genetiche della colorazione quaglia suggerite da Carefoot.