Vol. 3░ - VI.
I geni dellÆassenza
di pigmento melanico NEL PIUMAGGIO
|
|
Contrariamente a ci‗ che si potrebbe pensare di fronte
alle stupende varietÓ del piumaggio, esistono solo 2 pigmenti a determinarne il colore: il pigmento rosso e
quello nero, rispettivamente feomelanina e eumelanina.
Il
rosso per esprimersi necessita della presenza del gene s+,
il quale a sua volta, associato ad altri geni, dÓ tutte le gamme di colori
che variano dal frumento al mogano, passando per il fulvo, il dorato, il
marrone, il salmone.
Il
nero
per esprimersi in modo ottimale necessita della presenza di Ml
che dÓ il
nero integrale nel genotipo E,
e il gene Ml associato ad
altri geni crea a una serie di disegni secondari in cui il rosso Þ combinato
col nero, magari diluito in blu e grigio perla.
Il
bianco Þ dovuto allÆassenza di melanine. Le barbule
apigmentate sono cosý numerose da causare unÆimponente e diffusa riflessione
della luce, come accade per la neve, per il cotone o per la carta bianca.
Quando una piuma bianca Þ immersa nel balsamo del Canada, dotato di indice di
rifrazione
[1]
praticamente uguale a quello della cheratina, il bianco svanisce, e lo stesso
succede per un foglio di carta intinto in una sostanza oleosa. Siccome la riflessione dei raggi luminosi si verifica alla
superficie degli spazi aerei, un liquido con indice di
rifrazione 1,5 come il cresolo pu‗ spiazzare lÆaria dagli spazi
e rendere il materiale traslucido.
Il piumaggio bianco pu‗ accompagnarsi a vari gradi di
melanizzazione in altri tessuti: la melanina oculare pu‗ essere assente,
fortemente ridotta, oppure inalterata; la pelle e altri tessuti interni
possono essere o non essere interessati dallÆamelanosi.
Gli albini sono caratterizzati da un piumaggio bianco con
assenza o spiccata riduzione delle melanine oculari e possono essere
tirosinasi positivi o negativi. Non esistono esempi di amelanosi oculare che
non interessino anche la pigmentazione delle piume, sebbene la mutazione
occhio rosa pk sembri dotata di
maggior effetto sullÆocchio che sul piumaggio. Una sindrome ereditaria
caratterizzata da spontanea perdita di pigmentazione delle piume nel periodo
postnatale si accompagna a eliminazione della melanina epidermica e uveale.
Il piumaggio bianco Þ
economicamente desiderabile per lÆindustria alimentare della carne, poichÚ
non lascia residui di melanina nel follicolo dopo lÆasportazione delle
piume.
1. Bianco dominante
|
I - inhibitor |
Autosomico incompletamente dominante
Gruppo di associazione II -
cromosoma 2
Il
simbolo I
sta
a significare inibitore del pigmento,
in quanto un singolo gene Þ in grado di sopprimere la produzione dellÆeumelanina
con un lieve effetto sulla feomelanina. Il simbolo I fu usato per la prima volta da Hadley nel 1913
per
individuare gli effetti del gene scoperto da Bateson nel 1902.
Questa mutazione Þ caratteristica della Livorno bianca, ma Þ posseduta anche
dalla Cornish rossa orlo bianco e Giubileo, Polish fulva orlo bianco,
Wyandotte fulva orlo bianco, dai due Combattenti Inglesi Antico e Moderno pile,
dal Collo Nudo del Forez. Quei pulcini che da adulti saranno completamente
bianchi, quando nascono sono di colore giallo chiaro.
Il gene I pu‗ essere presente in certe razze e varietÓ bianco
recessivo come la Minorca bianca e la Dorking bianca, e proprio con genotipi I_I/c_c
si ottiene un bianco particolarmente puro.
Dire Livorno bianca significa bianco dominante. Tuttavia
pare che certi ceppi particolarmente candidi abbiano un colore di base E_E,
dove I allo stato omozigote va a braccetto con il barrato legato al
sesso - o col suo allele BSd - nonchÚ, in qualche
caso, col gene del blu e con quello dellÆargento.
Per nostra comoditÓ riportiamo nuovamente quanto esposto
nel II volume circa la sintesi melanica. I melanosomi vengono
prodotti attraverso il coordinamento di due momenti separati e geneticamente controllati: uno Þ responsabile della
sintesi e dellÆassemblamento della matrice
del premelanosoma
[2]
,
lÆaltro
coordina la formazione e la liberazione
del complesso tirosinasi-tirosina verso il premelanosoma.
I
premelanosomi originano nel reticolo endoplasmatico. La
tirosinasi Þ sintetizzata nel reticolo endoplasmatico granulare, Þ
organizzata e complessata nellÆapparato di Golgi, quindi viene trasportata
dalle vescicole alla matrice del melanosoma. Quando le vescicole aderiscono
alla membrana del premelanosoma, lÆenzima viene attivato, il polimero
melanico viene sintetizzato e attaccato alla matrice.
Pare che il meccanismo dÆazione della mutazione I
consista nella soppressione della formazione dei melanosomi.
Brumbaugh ha dimostrato che viene ridotta lÆincorporazione di 3-H-leucina
entro i melanosomi di genotipo I/I,
ma non in quelli con genotipo i+/i+,
ridotta incorporazione che Þ indicativa di una sintesi rallentata.
╚ presente un rallentamento
della formazione delle vescicole del Golgi entro le quali il
pigmento nero viene accumulato, per cui lÆeumelanina non ha dove venir
immagazzinata prima di passare ai cheratinociti. Il risultato finale Þ una
ridottissima produzione di pigmento.
LÆesame strutturale di piume in via di crescita indica
che in seguito al disturbo biochimico i melanociti della Livorno contengono
melanosomi di forma irregolare e irregolarmente assemblati. LÆipomelanosi,
derivando da unÆinibizione
della sintesi melanica, dÓ luogo a melanosomi irregolarmente affastellati e
in numero ridotto, nonchÚ a un disturbo del loro trasferimento ai
cheratinociti,
cui fa seguito una precoce morte del melanocita. Apparentemente il melanocita
cerca di proteggersi da questi organuli anomali e citotossici impacchettandoli
per essere degradati dagli autofagosomi, il che in ultima analisi porta alla
morte cellulare e alla riduzione di melanina nei cheratinociti.
╚ indispensabile un richiamo di
embriologia giÓ analizzato nel capitolo delle melanine (vol.II
-
XXVII).
I melanociti riconoscono una duplice origine embriologica:
º
lÆepitelio pigmentato della retina e del corpo ciliare,
lo strato pigmentato posteriore dellÆiride (o epitelio pigmentato dellÆiride),
forse anche le cellule pigmentate del pecten
[3]
derivano tutti quanti dallo strato esterno della coppa ottica
º
i melanoblasti che migreranno in tutti gli altri distretti
dellÆorganismo, inclusa la porzione uveale dellÆocchio
[4]
- composta da coroide, gran parte del corpo ciliare e superficie anteriore dellÆiride
(o endotelio dellÆiride) - provengono dalla cresta neurale.
LÆiride, cui spetta il ruolo maggiore nel caratterizzare
il colore dellÆocchio, pu‗ essere dotata di pigmento sia sulla faccia
anteriore che posteriore, le quali hanno origine embriologica diversa; pu‗
inoltre presentare dei melanociti che circondano i vasi sanguigni distribuiti
nel contesto dello stroma.
Questi dati si oppongono a ci‗ che si era sempre pensato,
e cioÞ che la sorgente dei melanociti fosse rappresentata da un singolo tipo
cellulare: quelle cellule che nascono dalla cresta neurale e che
successivamente prendono posto nei tessuti pigmentati. Le differenze
morfologiche e funzionali presenti nei diversi distretti sarebbero poi
scaturite da interazioni fra melanociti e ambiente.
Inoltre nel 1986 Boissy
osserv‗ che colture di cellule della cresta neurale contengono due tipi distinti di melanociti:
un tipo produce un pigmento uguale a quello delle piume che stanno
rigenerando, mentre lÆaltro contiene melanosomi simili a quelli della
coroide. Da ci‗ si pu‗ dedurre che pur avendo la stessa origine i
melanoblasti della cresta neurale si differenziano giÓ durante la vita
embrionale e che tale differenziazione non dipende dallÆambiente, bensý da
fattori di origine strettamente genetica.
Si pu‗ affermare che il gene I
Þ un elemento regolatore metabolico con azione ristretta alla cellula
pigmentaria.
Dal momento che il bianco dominante interferisce con la pigmentazione delle
piume e della coroide, ma non con lÆepitelio della retina e della faccia
posteriore dellÆiride, il gene I
agisce solo sui melanociti originati dalla cresta
neurale. Tuttavia, anche se lÆeumelanocita e il feomelanocita
del piumaggio hanno origine dalla cresta neurale, il secondo, o meglio lÆazione
feomelanica viene scarsamente inibita da I.
Dal momento che gli eumelanociti producono un maggior
numero di granuli con maggior quantitÓ di melanina rispetto ai feomelanociti
[5]
,
questo potrebbe spiegare perchÚ la feomelanizzazione sfugga maggiormente allÆinibizione
da parte di I. Si pu‗ quindi
giungere alla conclusione che il bianco dominante ha unÆazione dose-dipendente su ambedue i
melanociti: infatti un genotipo I/I inibisce maggiormente sia lÆeumelanina che la feomelanina
rispetto al genotipo I/i+.
Le cellule produttrici di feomelanina hanno unÆattivitÓ
pi¨ lenta, per cui in presenza di un solo gene I le vescicole possono venir formate per tempo, tanto da
permettere al pigmento di venir immagazzinato senza che il processo di
feomelanogenesi venga alterato.
I
Þ incompletamente dominante rispetto al suo allele i+
e gli effetti di I sul
fenotipo dipendono dalla sua azione epistatica sulla melanina, innanzitutto
sullÆeumelanina. I pulcini e gli adulti I/i+ sono bianchi con
striature nere, oppure sono grigiastro chiaro sia a carico nel piumino
![]() che delle piume di copertura. La quantitÓ di chiazze nere si riduce se Þ
presente il barrato legato al sesso B.
Sulla feomelanina la mutazione I
ha un effetto inibitore relativamente scarso ed Þ senza dubbio per questa
ragione che la Livorno bianca porta il gene E per mascherare il pigmento rosso e il gene S
per cancellare eventuali tracce di feomelanina nelle aree non nere.
che delle piume di copertura. La quantitÓ di chiazze nere si riduce se Þ
presente il barrato legato al sesso B.
Sulla feomelanina la mutazione I
ha un effetto inibitore relativamente scarso ed Þ senza dubbio per questa
ragione che la Livorno bianca porta il gene E per mascherare il pigmento rosso e il gene S
per cancellare eventuali tracce di feomelanina nelle aree non nere.
Le femmine pile
derivate da e+ hanno il petto salmone ![]() , mentre i maschi corrispondenti hanno mantellina,
piccole copritrici, triangolo dellÆala, dorso e sella color rosso oro,
talora mattone in certi punti, come le piccole copritrici
, mentre i maschi corrispondenti hanno mantellina,
piccole copritrici, triangolo dellÆala, dorso e sella color rosso oro,
talora mattone in certi punti, come le piccole copritrici
![]() -
- ![]() -
- ![]() -
- ![]() . Le aree bianche possono essere aumentate o ridotte da
una selezione volta ad alterare lÆestensione dellÆeumelanina. ╚
interessante il fatto che I
sia stato introdotto in alcuni ceppi fulvi in cui lÆeumelanina era
indesiderata nelle aree di restrizione columbia.
. Le aree bianche possono essere aumentate o ridotte da
una selezione volta ad alterare lÆestensione dellÆeumelanina. ╚
interessante il fatto che I
sia stato introdotto in alcuni ceppi fulvi in cui lÆeumelanina era
indesiderata nelle aree di restrizione columbia.

Fig. VI. 1
- Combattente inglese moderno nano
in
diverse varietÓ cromatiche fra cui quella pile
1.1. Pile oppure pyle
Il Giornale degli Allevatori n░ 7-8 del 1917 asseriva:
ôLÆetimologia del termine pile lÆignoro affatto, ma se questo fosse originato dal pile rovescio del face, potrebbe aversi in ci‗ la spiegazione, visto che la varietÓ in questione Þ precisamente il rovescio della medaglia, e nei caratteri del black red sta alla black red come la negativa di una lastra fotografica sta alla positiva stampata sulla carta. Tutte le parti scure sono bianche nella pile.ö
A me sembra pi¨ che ovvio
che lÆimpegno etimologico dellÆarticolista non fu per lui causa di stress
da ricerca.
Gli Inglesi usano scrivere pile, al contrario degli Statunitensi che preferiscono pyle.
Scorrendo la letteratura avicola si pu‗ notare che i due termini vengono
spesso usati indifferentemente. Dobbiamo tener presente che anche nei secoli
passati lÆortografia delle parole non sempre era corretta e che le lettere i
e y venivano usate senza una
cognizione precisa. Inoltre, quando la i
aveva un suono lungo come in eye, e
non breve come in it, spesso gli
amanuensi preferivano usare la lettera y.
Talora, invece, lo scrivere in un modo anzichÚ in un altro dipendeva dallÆestro
oppure da un senso estetico del tutto personali.
Sta di fatto che, dal punto di vista pratico, non
intercorre molta differenza tra pile
oppure pyle. LÆunico dato in
comune Þ che ambedue si pronunziano pi¨ o meno paial.
Per‗, come potremo arguire dallÆetimologia, la grafia corretta sarebbe pile.
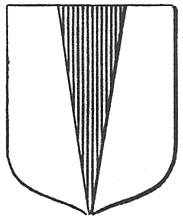
Fig. VI. 2 - Pila o Pile
A pagina 423 del Dizionario Araldico di Piero Guelfi Camaiani Þ riportata la definizione di pila, che in inglese Þ detta pile:
ôPezza
onorevole di seconda classe formata da un triangolo isoscele la base del quale
sta nella linea del capo ed il vertice nella punta. Quando il triangolo tocca
il centro del lato superiore dello scudo e la base Þ poggiata sulla linea
della punta chiamasi pila rovesciata. Se il triangolo (pila)
giunge sino al centro dello scudo dicesi pila scorciata.[...]ö.
La
pila Þ un segno frequente nellÆaraldica inglese, pi¨ raro
in quella italiana. Con un debole sforzo dellÆimmaginazione Þ facile
paragonare questa figura araldica alla sagoma disegnata dalle piccole
copritrici alari rosse contornate di bianco, oppure, con sforzo ancora minore,
al triangolo dellÆala che ha perso il contorno nero.
Vediamo ora lÆetimologia di pila.
Il giavellotto dei fanti romani era designato col nome di pilum di cui esistevano il tipo leggero e quello pesante ambedue
dotati di punta cuneiforme. QuestÆultimo era unÆarma grossolana che
ricordava la clava e che veniva scagliata prima di passare alla spada. Pilum
significava anche pestello, lÆutensile da cucina che serve a preparare un
buon pesto alla genovese. I pestelli pi¨ antichi usati dallÆuomo
preistorico per pestare, polverizzare, impastare o triturare le granaglie,
avevano la forma di un cuneo, erano di pietra e venivano manovrati entro
mortai formati da cavitÓ naturali della roccia. In latino, schiacciare,
pigiare, si dice pinso o piso
[6]
e il loro participio passato Þ pistum.
Spesso, per ragioni eufonetiche, la lettera s
viene sostituita dalla lettera l. LÆorigine
di pilum e di pile
Þ quindi lampante.
In sintesi, pile
si ricollega alla punta dellÆarma usata dalla fanteria romana - il pilum
- e originariamente questo vocabolo indicava il pestello che non era dotato di
estremitÓ rotonda come adesso, bensý cuneiforme come la punta della lancia.
Le piccole copritrici alari color rosso mattone contornate da bianco del
Combattente Inglese pile, oppure il suo triangolo dellÆala,
riproducono appunto una figura cuneiforme, usata anche in araldica.
1.2. Conflitti fra geni e allevatori
In sintesi: il gene I,
assente nel dorato che possiede il suo allele i+,
inibisce il nero comportandosi da epistatico rispetto a C+, e riduce il rosso. Allo stato omozigote
cancella
integralmente il nero e attenua molto il rosso; se Þ eterozigote
ha
scarsa azione sul rosso e pu‗ lasciar apparire qualche rara traccia di nero.
Gli allevatori lo chiamano anche Bianco
Livorno, perchÚ Þ stato osservato per la prima volta in questa razza. Si
sono osservati soggetti bianchi omozigoti che presentano alcune macchie rosse
e questo fatto denuncia che il gene I non sempre prevale sulla feomelanina.
Ogni allevatore di Cornish Þ a conoscenza del fatto che un singolo gene I
Þ in grado di trasformare un soggetto scuro nella varietÓ Giubileo, in cui
Þ frequente riscontrare delle tracce di nero, specie a carico della
mantellina e della coda dei maschi. LÆincrocio di un Giubileo con un
Giubileo Þ disapprovato dagli allevatori, poichÚ essi dicono che questo modo
di procedere porta a dei soggetti slavati, come effettivamente accade. Gli
slavati, per‗, servono egregiamente per essere accoppiati con quelli scuri al
fine di ottenere un giubilante Giubileo pari al 100%.
Lo stesso accade per la Wyandotte oro orlo nero: introducendovi il gene del bianco
dominante si ottiene una Wyandotte fulva orlo bianco, e procedendo nei
reincroci si ottengono soggetti slavati che corrispondono a uno stato
omozigote per I. A risolvere
questo problema viene in soccorso il gene del blu che in doppia dose trasforma
la varietÓ oro orlo nero in oro orlo bianco
![]() ,
in cui la presenza di Bl Þ
tradita sia dallÆintensitÓ maggiore del rosso al centro della piuma, sia da
residui di blu al capo e alla coda. Questa soluzione Þ stata adottata anche
nella Cornish rossa orlo bianco, che Þ una varietÓ diversa dalla Giubileo in
quanto, oltre ad avere la feomelanina integra, Þ una mono-orlata.
,
in cui la presenza di Bl Þ
tradita sia dallÆintensitÓ maggiore del rosso al centro della piuma, sia da
residui di blu al capo e alla coda. Questa soluzione Þ stata adottata anche
nella Cornish rossa orlo bianco, che Þ una varietÓ diversa dalla Giubileo in
quanto, oltre ad avere la feomelanina integra, Þ una mono-orlata.
La Sebright camoscio ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() - che Þ una fulva orlo bianco
come la
Polish camoscio
e la Padovana
camoscio - rientra nella stessa situazione genetica della
Cornish Giubileo. Nella Polish e nella Padovana Þ pi¨ facile raggiungere
questo risultato rispetto alla Wyandotte, in quanto alcuni ceppi di Polish
sono dotati di bianco dominante, mentre per lo pi¨ la Wyandotte Þ
caratterizzata da bianco recessivo per cui il gene del bianco dominante Þ da
prendere a prestito da unÆaltra razza.
- che Þ una fulva orlo bianco
come la
Polish camoscio
e la Padovana
camoscio - rientra nella stessa situazione genetica della
Cornish Giubileo. Nella Polish e nella Padovana Þ pi¨ facile raggiungere
questo risultato rispetto alla Wyandotte, in quanto alcuni ceppi di Polish
sono dotati di bianco dominante, mentre per lo pi¨ la Wyandotte Þ
caratterizzata da bianco recessivo per cui il gene del bianco dominante Þ da
prendere a prestito da unÆaltra razza.

Fig. VI. 3 - Padovana camoscio
Il
ciuffo bianco della Polish Þ stato attribuito da Somes a un gene
incompletamente dominante capace di determinare un ciuffo ampiamente
depigmentato su uno sfondo nero integrale oppure blu. Tuttavia pare che
questo gene non sia ancora stato analizzato in modo soddisfacente. Nonostante
ci‗, si pu‗ senzÆaltro affermare che esso comporta unÆinterruzione del
processo di melanizzazione confinato al ciuffo. UnÆaltra ipotesi
suggerirebbe la mancata migrazione dei melanociti in questo distretto. A
questo proposito si rimanda al problema del mosaicismo e degli albinismi
localizzati (vol.II
- XXI, 2.1.). Tuttavia, a mio giudizio e in base alla mia limitata
esperienza, nella Polish nera ciuffo bianco deve esistere qualche problema a
carico della popolazione melanocitaria: non solo le piume del mantello adulto
sono spesso melanizzate in modo incompleto e bizzarro, in quanto anche nei
pulcini si possono notare vaste aree non eumelanizzate ![]() -
- ![]() -
- ![]() -
- ![]() a differenza di quanto accade per varietÓ nere di altre razze
a differenza di quanto accade per varietÓ nere di altre razze
![]() -
- ![]() -
- ![]() -
- ![]() .
.
In occasione di Ornithophilia 1994 tenutasi a Utrecht era esposta una nuova colorazione di Barbuta di Watermaal: Quaglia limone bianco, wit citroenkwartel. Rivolsi subito una domanda al Presidente del Club delle Barbute Belghe, il Professor Hans Heemskerk, per sapere se per caso la Barbuta di Watermaal fosse dotata di bianco dominante oppure di bianco recessivo. Egli mi disse che le Barbute Belghe sono sfornite di bianco dominante, ma non poteva mettere la mano sul fuoco. Dopo pochi minuti il disponibilissimo Hans mi diede la possibilitÓ di uno scambio di idee con un allevatore belga del quale non annotai il nome e che escluse in modo categorico la possibilitÓ di bianco dominante nelle nane belghe. Io ebbi un bel ribattere che il bianco recessivo cancella ogni traccia di rosso dal mantello, ma lÆinterlocutore fu irremovibile, anche quanto prospettai la possibilitÓ che fosse stato introdotto un bianco dominante come nella Wyandotte fulva orlo bianco.
Pi¨ ci penso e pi¨ son convinto che egli
avesse torto, in quanto il bianco recessivo ubbidisce alla legge del tutto o nulla,
Þ impietoso e non accetta compromessi, contrariamente al bianco dominante il
quale, anche se omozigote, per alcune generazioni pu‗ lasciar trapiumare
la feomelanina. Queste affermazioni non devono essere prese come assiomi, in
quanto vedremo tra poco che piccole deroghe sono ammesse.
I soggetti quaglia
limone bianco erano quasi totalmente bianchi, eccetto una tenue sfumatura
beige lÓ dove Þ normalmente presente feomelanina in una Barbuta di Watermaal
ben eumelanizzata. Se ha ricevuto anche lÆaggettivo limone, Þ da presumere
la presenza di ig a diluire la
feomelanina. Io escludo il bianco recessivo e senza dubbio ci sta a pennello
il bianco dominante in singola dose, in quanto il rimanente effetto schiarente
sulla feomelanina Þ svolto da ig.
Naturalmente mi ero prefisso di chiedere notizie allÆallevatore olandese.
Ma ci ha pensato Aad Rijs, ottimo
allevatore di Barbute di Grubbe, incaricato da Heemskerk a chiarire il mistero
che veniva cosý dipanato da una sua lettera in data 23
gennaio
1995:
|
Dear
mister Corti, |
Caro
signor Corti, |
|
In the past I received
information directly from the originator of this beautiful colour. He
told me that he found his first birds in this colour in a hatch of
imported eggs from unknown Belgium parents. Through the process of
selecting he developed a strain in this colour. |
In passato ho ricevuto dirette
informazioni da chi ha dato origine a questa bella colorazione: mi ha
detto che ha trovato i primi soggetti di questo colore in una schiusa
di uova importate e provenienti da genitori Belgi
sconosciuti. Con la selezione ha svilup¡pato una linea dotata di
questo colore. |
|
Due to the fact that his
birds had always a perfect type, a lot of breeders crossed the
wit-citroenkwartels with normal quails. The outcome in the F1 crossbred was always white-quail (quaille
blanc). In other words the white from the wit-citroenkwartel dominated
the black from the normal quail, which proves that the dominant gene
for white is involved. Furthermore we learned from this cross breeding
that the lemon/citron colour was dominated by the ônormalö gold
colour from the quails. This proves that also the recessive gene for
lemon/citron is involved. |
Siccome questi soggetti
mantenevano sempre un tipo perfetto, diversi allevatori hanno
incrociato i wit-citroen-kwartel con la normale colorazione quaglia.
La progenie F1
di
questo incrocio risultava sempre bianco-quaglia. In altre parole, il
bianco del wit-citroenkwartel era dominante sul nero della quaglia
normale, e questo Þ la riprova che Þ coinvolto il gene del bianco
dominante. Inoltre, da questo tipo di incrocio abbiamo imparato che il
limone viene dominato dal normale
oro che caratterizza la colora¡zione quaglia. Questo dimostra che Þ
coinvolto an¡che il gene recessivo per il limone. |
|
You state in your letter
that you spoke with a Belgium breeder who told you that there is no
gene for dominant white in Belgium bantams. I can assure you that this
statement is absolutely wrong! |
Nella sua lettera lei afferma che ha parlato con un
allevatore belga il quale le ha detto che non esiste il gene del
bianco dominante nelle Nane Belghe. Posso assicurarle che questÆaffermazione
Þ del tutto errata. |
|
We ourselves had a white
hen which we crossed with a true breeding cuckoo male. We expected to
get better offspring with no gold in the feathers. Instead of that we
ended up with only white youngsters due to the fact that the white
mother carried the dominant white gene. We crossed the white F1 together and ended up with 7 different
colours, which proved again that the dominant white gene was involved.
Amongst them where quails, blue quails, white quails, self-blue,
cuckoo, blue-cuckoo and quail with cuckoo markings. |
Io stesso avevo una gallina bianca che ho incro¡ciato
con un maschio cuc¨ puro. Mi aspettavo la miglior discendenza con
assenza di oro nelle penne. Invece di ottenere ci‗, mi sono ritrovato
solamente con dei soggetti pi¨ giovani di colore bianco, in quanto la
madre bianca era portatrice del bianco dominante. Ho incrociato i
soggetti bianchi di F1
tra
loro e ne sono usciti 7 diversi colori, confermando di nuovo il
coinvolgimento del gene del bianco dominante, in quanto i colori
erano: quaglia, quaglia blu, quaglia bianco, blu, cuc¨, blu-cuc¨ e
quaglia con disegno cuc¨. |
|
Furthermore the colour
yellow-white mottled is standardised in Holland and Belgium. In fact
this colour is strictly related to porcelain with the only distinction
that that black spots on the feathers of the porcelain are exchanged
by white. Crossing both colours together if bred true, gives you only
yellow-white mottled in the F1 generation. Crossing the F1 together gives you 25% porcelain and 75% yellow-white
mottled. Again is proven that the gene for dominant white is involved. |
Inoltre, la colorazione millefiori giallo-bianco Þ
riconosciuta dallo standard sia in Olanda che in Belgio. In effetti,
questo colore Þ strettamente correlato al millefiori, fatta eccezione
per la chiazza nera del millefiori che viene sostituita dal bianco. Se
le due colorazioni sono pure e vengono tra loro incrociate, in
F1
otterremo solo soggetti millefiori giallo-bianco. Incrociando la
progenie F1
si
ottiene un 25% millefiori e un 75% millefiori
giallo-bianco. Ecco di nuovo la dimostrazione del coinvolgimento del
gene del bianco dominante. |
|
I hope that this letter
has been of help to you and I wish you much success with the breeding
of your Belgium birds. |
Spero che questa lettera le sia di aiuto e le au¡guro
molto successo nellÆallevamento delle sue Bar¡bute Belghe. |
Kind regards. Aad F.J.Rijs - Ingen - Holland
2. Marrone grigiastro
|
ID
- dun |
Autosomico incompletamente dominante, allele di
I rispetto al quale Þ recessivo
Gruppo di associazione II -
cromosoma 2
In
seguito allÆosservazione fatta da Hollander (1981) su
una femmina marrone grigiastra nata da un gallo dÆarena dal mantello
blu-rosso insolitamente chiaro e da una gallina Sumatra nera, Þ stata
proposta lÆesistenza di questo allele di I,
anchÆesso attivo prevalentemente sullÆeumelanina negli eterozigoti (1987).
Il fenotipo sembrava quello della mutazione pk - pink eye - eccetto a
livello della retina, che non era rosa.
LÆeffetto eterozigote si traduce in una colorazione
marrone grigiastra, mentre lo stato omozigote determina un mantello
biancastro. Inoltre, mentre gli eterozigoti I/i+ presentano spesso
macchie nere, gli eterozigoti I/ID presentano macchie in numero relativamente scarso e
sono di colore dun, cioÞ
marrone grigiastro. Non sono state fornite sicure indicazioni sulle
relazioni tra I e ID; tuttavia il primo pare dominante sul secondo.
3. Il locus C+ - color
Un dato Þ certo: questo locus
possiede diversi alleli. Il fenotipo noto come bianco recessivo c
Þ il pi¨ frequente tra le varietÓ bianche non dovute al bianco dominante e
sono stati dimostrati i suoi effetti epistatici su altri geni responsabili del
colore.
Essendo molto probabile che il gene C+ sia il gene strutturale della tirosinasi contenuta nelle vescicole di
Golgi, qualsiasi soggetto colorato possiede almeno un gene C+.
Due alleli, cre e
ca,
non presentano attivitÓ tirosinasica.
Ecco i diversi alleli che
possono occupare questo locus e le loro sigle:
c - bianco recessivo
cre
- red eye -
bianco
occhio rosso
ca
- albinismo
recessivo o autosomico
con
il seguente rapporto di dominanza:
C+
> c > cre
> ca
Prima di procedere vale la pena di dare uno sguardo alla
tabella riassuntiva in cui compare anche lÆalbinismo imperfetto legato al
sesso di cui riparleremo nonostante sia giÓ stato trattato in vol.III
- II capitolo.
| Geni
che causano un piumaggio bianco e fenotipi degli omozigoti |
||||
| nome | simbolo | dominanza | piumaggio | occhio |
| Bianco dominante | I | incompleta | bianco | baio |
| Marrone grigiastro - dun | ID | incompleta | biancastro | pigmentato |
| Bianco recessivo | c | recessivo | bianco | per lo pi¨ baio (1) |
| Bianco occhio rosso | cre | recessivo | bianco | rosso scuro |
| Albinismo recessivo | ca | recessivo | bianco | rosa |
| Albinismo imperfetto | sal | recessivo | in parte pigmentato (2) | rosso chiaro |
| (1) -
pu‗ essere in parte o totalmente marrone con appropriati genotipi (2) - il piumaggio nero diventa marrone grigiastro e quello rosso diventa rosso chiaro |
||||
I colori del piumino dei pulcini
dotati di uno dei tre mutanti c
-
cre
- ca
allo stato omozigote indicano che tutti i genotipi, tranne ca/ca,
sono in grado di produrre un poÆ di eumelanina.
Il piumino del bianco recessivo c Þ molto variabile: pu‗ essere chiaro ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() , oppure pu‗ essere
scuro quasi come nei maschi di Plymouth Rock barrata. In altri casi i pulcini c/c
hanno un piumino amelanotico. Non Þ ancora stato chiarito quali sono i
genotipi che fanno da sfondo a queste variazioni cromatiche del piumino e si
pu‗ solo affermare che il gene E
Þ presente quando il piumino si presenta grigiastro. LÆintensitÓ del
grigio sembra dovuta a geni modificatori, anche se le femmine in media si
presentano pi¨ scure dei maschi. Le conoscenze relative alle colorazioni
rosso scuro o grigio giallastro sporco sono scarse, anche se in due
popolazioni studiate da Smyth si poteva escludere la presenza di E.
Nonostante non sia ancora stato isolato un genotipo associato al piumino
bianco, in alcuni casi pu‗ essere definito come la combinazione dei geni
ipostatici I ed E. Il piumino degli omozigoti cre/cre
pu‗ essere sia bianco che leggermente grigio.
, oppure pu‗ essere
scuro quasi come nei maschi di Plymouth Rock barrata. In altri casi i pulcini c/c
hanno un piumino amelanotico. Non Þ ancora stato chiarito quali sono i
genotipi che fanno da sfondo a queste variazioni cromatiche del piumino e si
pu‗ solo affermare che il gene E
Þ presente quando il piumino si presenta grigiastro. LÆintensitÓ del
grigio sembra dovuta a geni modificatori, anche se le femmine in media si
presentano pi¨ scure dei maschi. Le conoscenze relative alle colorazioni
rosso scuro o grigio giallastro sporco sono scarse, anche se in due
popolazioni studiate da Smyth si poteva escludere la presenza di E.
Nonostante non sia ancora stato isolato un genotipo associato al piumino
bianco, in alcuni casi pu‗ essere definito come la combinazione dei geni
ipostatici I ed E. Il piumino degli omozigoti cre/cre
pu‗ essere sia bianco che leggermente grigio.
Tutti e tre
i piumaggi adulti sono bianchi anche se il piumino
alla nascita era scuro. Quei soggetti dotati dellÆallele c
che si presentano leggermente grigi derivano da incroci tra ceppi dei
quali uno era portatore di intensificatori dellÆeumelanina.
Come giÓ detto, studi ultrastrutturali e citochimici
hanno dimostrato che i melanociti delle piume dei mutanti cre e ca sono privi di tirosinasi. Si Þ visto che i melanociti
tentano di fabbricare i melanosomi, che tuttavia sono incompleti e poco
numerosi, tanto da causare lÆipertrofia dellÆapparato di Golgi. Inoltre
sono presenti numerose vescicole di premelanosomi contenenti un materiale che
Þ stato definito matrice tipo lattice.
Al momento attuale non si pu‗
affermare con sicurezza se gli alleli di C+ rappresentano delle
mutazioni del gene strutturale della tirosinasi oppure se esercitano il loro
effetto sullÆenzima regolandone la funzione. Ambedue i geni cre e ca producono una molecola
simile alla tirosinasi dal punto di vista sia elettroforetico che
immunologico, sprovvista tuttavia di attivitÓ enzimatica. Ci‗ suggerirebbe lÆipotesi
secondo cui questi alleli di c
rappresentano una sostituzione aminoacidica in un sito attivo, anche se per
queste mutazioni non si possono escludere delle modificazioni
post-traduzionali. In ogni caso, sia per c
che per cre
e per ca
si pu‗ affermare che si tratta di geni
fiacchi, geni che fanno
acqua, leaky genes per usare un
termine tanto caro agli anglofoni.
4. Bianco recessivo
|
c
- recessive white |
Autosomico recessivo - allele di C+
Gruppo di associazione
sconosciuto
Tutto
quanto riguarda lÆallele c lÆabbiamo
testÚ esaminato parlando del locus del colore. Pertanto, salvo piccole
aggiunte, quanto segue Þ un riassunto di ci‗ che abbiamo appena visto.
Il bianco recessivo Þ responsabile del bianco
maggiormente diffuso tra quei polli ornamentali che non sono dotati di bianco
dominante, e in senso assoluto pare sia il gene maggiormente responsabile del
bianco. La sua azione nellÆinibire il rosso Þ molto pi¨ efficace rispetto
a quella del gene del bianco dominante, in quanto il suo meccanismo dÆazione
impedisce le reazioni chimiche necessarie alla formazione dei materiali di
base con cui vengono allestite le vescicole del reticolo endoplasmatico in cui
trovano posto i melanosomi, e bisogna puntualizzare il fatto che molto
probabilmente C+ Þ il gene strutturale della
tirosinasi. Pertanto viene alterata la sintesi di ambedue i tipi di pigmento.
E si badi bene, non si tratta di un semplice rallentamento della formazione
delle vescicole come nel bianco dominante, ma di unÆinibizione.
Talora lÆazione di c
Þ deficitaria, per cui si possono riscontrare delle striature di
pigmento. Spesso nelle
remiganti secondarie della Wyandotte si riscontrano delle aree
grigiastre, in aggiunta alle quali non Þ raro trovare alcune tracce dei due
pigmenti, specialmente nella mantellina dei maschi.
In un certo senso il bianco recessivo potrebbe essere
considerato un albinismo, ma Þ corretto riservargli il termine di albinismo delle piume, in quanto lÆocchio, i tarsi e la cute non sono
interessati.
Il
piumino del bianco recessivo Þ molto variabile: pu‗ essere
chiaro ![]() -
- ![]() -
- ![]() -
- ![]() -
- ![]() , oppure pu‗ essere scuro quasi come quello dei maschi di Plymouth Rock
barrata. In altri casi i pulcini c/c
hanno un piumino amelanotico. Quando il piumino tende al grigio Þ
presente il gene E, nel qual
caso lÆintensitÓ del grigio dipende da agenti modificatori che rendono le
femmine pi¨ scure dei maschi.
, oppure pu‗ essere scuro quasi come quello dei maschi di Plymouth Rock
barrata. In altri casi i pulcini c/c
hanno un piumino amelanotico. Quando il piumino tende al grigio Þ
presente il gene E, nel qual
caso lÆintensitÓ del grigio dipende da agenti modificatori che rendono le
femmine pi¨ scure dei maschi.
Quando il piumino Þ perfettamente bianco, ci‗ Þ stato
attribuito alla combinazione con il gene I
e con il gene E, che sono
ipostatici.
Il bianco recessivo pu‗ presentare alcune pigmentazioni,
abitualmente grigio chiaro, che derivano da incroci con ceppi commerciali i
quali tendono a portare con sÚ dei fattori di forte intensificazione dellÆeumelanina.
Altri ceppi sono dotati di forte eumelanizzazione delle gambe e dellÆocchio,
tanto da somigliare a dei bianchi dominanti eterozigoti su uno sfondo E.
Accoppiando
tra loro soggetti puri per il bianco recessivo talora
compaiono dei discendenti colorati. Questo apparente mistero pu‗
essere spiegato attraverso mutazioni insorte a livello del locus occupato da c.
La nuova mutazione non causa nulla al soggetto che ne Þ stato colpito, egli
rimane bianco, ma incrociando soggetti che sono andati incontro a mutazioni in
punti differenti di c,
compariranno soggetti colorati. Vediamo cosa accade.
Soggetto
bianco non mutato
---
A ------------ B ---
---
A ------------ B ---
Le linee spezzate e le lettere interposte indicano il
locus c. Nel locus occupato da
c possiamo indicare due
diversi frammenti di DNA con le lettere A e B
Supponiamo ora di avere un
genitore che Þ andato incontro a una mutazione nel frammento A di uno dei due cromosomi e
indichiamo tale mutazione con a:
Soggetto
mutato ma bianco
---
a ------------ B ---
---
A ------------ B ---
Il soggetto conserverÓ il suo fenotipo bianco, ma lo
stesso non accadrÓ alla sua discendenza.
Infatti, incrociato con un
soggetto non mutante, si otterrÓ un 50% di soggetti colorati:
Gameti:
---
a ------------ B --- x
---
A ------------ B ---
con lÆaccoppiamento il corredo
genico sarÓ rappresentato da:
|
--- a ------------ B --- |
aABB = soggetto colorato |
|
--- A ------------ B --- |

Complete
association between a retroviral insertion
in the tyrosinase gene
and the recessive white mutation in chickens
2006
4.1. Applicazioni pratiche del bianco recessivo
Il bianco recessivo omozigote
dÓ una soppressione completa sia del rosso che del nero. Per cui non sarÓ mai possibile ottenere il pile.
QuestÆaffermazione, naturalmente, non Þ valida in assoluto, in quanto
abbiamo visto che nella Wyandotte sono spesso presenti residui dei due
pigmenti. Per‗ Þ inutile intraprendere lÆavventura di voler ottenere dei
soggetti pile incrociando un bianco recessivo col suo omologo colorato: i
risultati sarebbero deludenti, anzi nulli.
Dagli incroci tra una varietÓ bianco recessivo col suo
omologo colorato in cui Þ presente il gene C+,
tutti i discendenti saranno colorati a meno che C+
fosse eterozigote nel genitore pigmentato. Pertanto lÆaffermazione che c
Þ completamente recessivo rispetto a C+
Þ senza dubbio valida.
Si afferma, giustamente, che il bianco recessivo in
singola dose lascia inalterata la colorazione di base, ma Carefoot ha potuto
dimostrare che la
presenza di un solo gene c
pu‗ essere scoperta in linee chiuse dotate di una precisa colorazione, e
precisamente in ceppi
di Wyandotte perniciata deputata alla produzione di maschi, nella quale i
maschi portatori di un solo gene c
hanno una
mantellina
limone brillante ![]() -
- ![]() , tantÆÞ che incrociando un
gruppo di soggetti con mantellina molto chiara scaturiscono automaticamente
soggetti per ╝ bianchi, per ╝ rossi e metÓ sono chiari.
[7]
La produzione in modo regolare di soggetti bianchi Þ inevitabile e deve
essere per forza accettata se si vogliono ottenere sia soggetti a mantellina
chiara che soggetti con spalle da
pettirosso. ╚ logico che esistono, e sempre esisteranno, variazioni a
carico della diluizione nel contesto di soggetti a mantellina chiara per
variazioni nel corredo di geni modificatori. Sfortunatamente questo effetto
diluente esercitato da una singola dose di c
non interessa solamente i pigmenti rosso e arancio. Interessa anche il nero,
la cui intensitÓ si riduce, rendendo spesso bianco lÆiporachide dei maschi
chiari, e anche le loro remiganti primarie presentano abitualmente un poÆ di
bianco che viene spesso penalizzato dai giudici in modo severo, anche se cosý
non dovrebbe essere se si tiene conto del meccanismo genetico che conduce alla
tonalitÓ chiara.
, tantÆÞ che incrociando un
gruppo di soggetti con mantellina molto chiara scaturiscono automaticamente
soggetti per ╝ bianchi, per ╝ rossi e metÓ sono chiari.
[7]
La produzione in modo regolare di soggetti bianchi Þ inevitabile e deve
essere per forza accettata se si vogliono ottenere sia soggetti a mantellina
chiara che soggetti con spalle da
pettirosso. ╚ logico che esistono, e sempre esisteranno, variazioni a
carico della diluizione nel contesto di soggetti a mantellina chiara per
variazioni nel corredo di geni modificatori. Sfortunatamente questo effetto
diluente esercitato da una singola dose di c
non interessa solamente i pigmenti rosso e arancio. Interessa anche il nero,
la cui intensitÓ si riduce, rendendo spesso bianco lÆiporachide dei maschi
chiari, e anche le loro remiganti primarie presentano abitualmente un poÆ di
bianco che viene spesso penalizzato dai giudici in modo severo, anche se cosý
non dovrebbe essere se si tiene conto del meccanismo genetico che conduce alla
tonalitÓ chiara.
Il bianco recessivo Þ presente in Cocincina, Cocincina
nana, Dorking, Nagasaki ![]() -
- ![]() -
- ![]() ,
Langshan, Orpington, Moroseta
,
Langshan, Orpington, Moroseta ![]() -
- ![]() ,
Sultano
,
Sultano ![]() -
- ![]() ,
Wyandotte, Bresse, Gatinaise, Plymouth Rock, Minorca, Jersey Giants e altre razze ancora. Questi bianchi hanno talora tendenza a mostrare riflessi paglia.
,
Wyandotte, Bresse, Gatinaise, Plymouth Rock, Minorca, Jersey Giants e altre razze ancora. Questi bianchi hanno talora tendenza a mostrare riflessi paglia.

Fig. VI. 4 - Silky bianca con barba e senza barba insieme alle varietÓ nera e oro
In certi ceppi di Bresse bianca, che danno regolarmente prole grigia,
il piumaggio, oltre a eventuali riflessi paglia, presenta il barrato
autosomico della Bresse grigia. Il bianco recessivo c/c
non riesce a inibire completamente la sintesi di melanina generata dal barrato
autosomico. Questa comparsa di Bresse grigie a partire da un ceppo bianco Þ
un enigma genetico. Dams ravvisa lÆesistenza di uno o pi¨ geni che
permettono lÆespressione del barrato a seconda delle combinazioni geniche.
5. Albinismo recessivo autosomico
|
ca
- autosomal albinism |
Autosomico recessivo - allele di C+
Gruppo di associazione
sconosciuto
Questo
allele comporta la scomparsa di qualsiasi pigmento in qualsiasi distretto dellÆorganismo.
Nel 1933 Warren scoprý un albinismo
totale caratterizzato da soggetti bianchi con occhi rosso brillante dovuti allÆassenza
di pigmenti che nellÆiride del soggetto normale mascherano il rosso del
sangue capillare. Warren descrisse questo fenotipo in un ceppo di Wyandotte
bianca.
Osservando certi ceppi di Wyandotte nana bianca, Carefoot
Þ dellÆavviso che questo gene indesiderato, o un suo corrispettivo, sia
ancora presente tra noi. Spesso vengono esposti, nonchÚ tollerati, polli
dalla pelle bianca e dallÆocchio rosso, senza obbedire allo standard che
richiede un occhio baio. La ragione pi¨ probabile di tutto ci‗ sta nel fatto
che lÆalbinismo tende a dare un piumaggio pi¨ bianco del solito. Questa
mutazione, sotto certi versi, riceverebbe senzÆaltro il benvenuto qualora
riuscisse a contaminare certi ceppi di Orpington bianca! Il meccanismo dÆazione
di questa mutazione dovrebbe essere del tutto simile a quello dellÆallele c.
Gli effetti negativi dellÆalbinismo
recessivo verranno tra poco descritti a proposito delle pleiotropie
ipomelaniche.
6. Albinismo imperfetto
|
sal
-
imperfect albinism |
Legato al sesso, recessivo, allele di s+ e di S
Gruppo di associazione V -
cromosoma Z
Ci
siamo giÓ soffermati su questa mutazione in vol.III - II.
Questo albinismo fu denominato imperfetto in quanto non era in grado di
eliminare completamente il pigmento nella Plymouth Rock barrata in cui fu
descritto per la prima volta. Tali soggetti erano dotati di una barratura
fantasma. I melanociti occupano la
loro normale sede nei follicoli, ma contengono uno scarso numero di
premelanosomi morfologicamente normali, mentre numerosi sono quelli
anormali. Pare che il difetto enzimatico consista in un trasporto inefficiente
della tirosinasi e della fosfatasi acida ai premelanosomi.
Su sfondo E
il fenotipo di questo albinismo sfocia in un marrone grigiastro, sia nel pulcino che nellÆadulto.
Il colore dellÆocchio va da un rosa chiaro a un rossastro scuro, a seconda
del colore di base. Gli occhi rosa pi¨ chiari compaiono in presenza di I,
mentre occhi pi¨ scuri, che si scuriscono ulteriormente col passare del
tempo, si riscontrano nei soggetti pigmentati normalmente.
Gene raro, Þ il pi¨ recessivo della serie allelica S.
Pare che questa mutazione non sia presente nelle razze francesi.
7. PLEIOTROPIE IPOMELANICHE
Deleteri effetti pleiotropici
sono stati correlati a parecchi geni che inibiscono in modo spiccato la
melanizzazione in numerose specie animali, compreso lÆuomo e altri
mammiferi.
Nei polli lÆesempio migliore Þ fornito dal gene ca
dellÆalbinismo recessivo, associato a scarsa capacitÓ visiva con fotofobia.
Il genotipo omozigote per ca comporta altri effetti
pleiotropici statisticamente significativi i quali coinvolgono lo sviluppo
dellÆembrione e del pulcino nel primo periodo postnatale. Tali effetti sono:
piumino pi¨ corto, incidenza maggiore della protrusione del sacco del tuorlo,
emorragie e infiammazioni nel sottocutaneo, incremento della mortalitÓ sia
embrionale tardiva che postnatale sino allÆottava settimana, ridotta
velocitÓ di crescita postnatale. Si pu‗ agevolmente dedurre che lÆalbinismo
autosomico intacca certi meccanismi fondamentali che regolano lo sviluppo.
Anche lÆallele c possiede un piccolo ma
consistente effetto depressivo sulla velocitÓ di crescita, cosa che non
accade con I.
Anche il gene dellÆalbinismo imperfetto sal possiede effetti sulla crescita
quasi sovrapponibili a quelli determinati da ca,
e questo desta un poÆ di meraviglia poichÚ i due geni agiscono in modo
differente sulla melanizzazione: il primo Þ tirosinasi positivo, lÆaltro Þ
negativo.
8. AMELANOSI SPONTANEA POSTNATALE
Esistono soggetti normalmente
pigmentati che a un certo punto sviluppano un piumaggio parzialmente o
totalmente amelanotico. Talora sono in causa neoplasie e disordini endocrini,
come abbiamo detto nel capitolo del mosaicismo (vol.II
- XXI-2.1.), mentre numerosi casi hanno suggerito lÆipotesi di
una causa sconosciuta su base genetica.
QuestÆanomalia compare tra le 5 e le 20 settimane di vita.
Pare si verifichi qualcosa di simile a quanto accade nella vitiligine umana. LÆincapacitÓ a
melanizzare le piume si associa a unÆiperattivitÓ melanica. Un dato interessante Þ che lÆincidenza
dellÆamelanosi Þ maggiore nel piumino di genotipo eb/eb.
I melanosomi anormali conducono alla formazione di autofagosomi ed eventualmente alla morte cellulare. Questo
processo Þ seguito da una risposta immunitaria volta a eliminare i
melanociti. In seguito i melanoblasti migrano dalle loro riserve dermiche
verso la nuova piuma in via di crescita poichÚ i melanoblasti indifferenziati
non vengono distrutti dalle cellule immunitarie. Questa affermazione Þ
dimostrata da periodi di temporanea
rimelanizzazione che si verificano nelle femmine durante la muta
successiva al periodo depositivo.
Questi tipi di amelanosi interessano anche lÆuvea e il pigmento cutaneo.
In circa il 50% dei soggetti con estese aree di amelanosi del
piumaggio si instaura una malattia corioretinica su base immunitaria che porta
a cecitÓ (vedi XVII-5.5.). Tale corioretinite su base immune Þ innescata dalle reazioni
infiammatorie associate alla distruzione dei melanociti della coroide. ╚
presente un 3-4% di tiroiditi
autoimmuni e un difetto delle piume tipo alopecia, anchÆessa legata a una
risposta immunitaria.
LÆereditarietÓ coinvolge numerosi geni, oltre ad alcuni geni
modificatori. LÆespressione del difetto posseduto dalla cellula pigmentaria
Þ variabile e lÆanalisi genetica Þ complicata dalla necessitÓ di
ricorrere al microscopio elettronico per una corretta classificazione. Oltre a
fattori genetici, paiono implicati fattori ambientali che alterano lÆattivitÓ
dei primi.
Infatti nella Castigliana nana nera sono state riscontrate
piume nere con apice sfumato fino al bianco e pare che questo accada nei
soggetti pi¨ deboli o che hanno sofferto di qualche patologia durante la
crescita (Fabrizio Focardi, 1994,
comunicazione personale).
Nella classica varietÓ nera della Spagnola faccia bianca
le piume bianche sono limitate allÆVIII-IX-X remigante
primaria e scompaiono completamente con la prima muta. Nella discendenza di
Spagnola x
Minorca e di Spagnola x Java, le macchie bianche
limitate allÆapice delle penne erano presenti nelle timoniere e nelle
remiganti e sono ricomparse anche dopo la muta (Luciano Tona, 1994,
comunicazione personale).
L'impareggiabile
olandese Elly Vogelaar![]() all'inizio di novembre del 2012 Þ entrata in contatto con Hein van Grouw
(Life Sciences department - Natural History Museum - Tring - Hertfordshire - UK)
il quale si dedica alla ricerca relativa all'amelanosi negli uccelli, quell'amelanosi
che pu‗ fare la sua comparsa anche oltre le 20 settimane di vita, e magari
indipendentemente da fattori genetici. Hein van Grouw ha comunicato a Elly
quanto segue, che a mio avviso Þ di estremo interesse.
all'inizio di novembre del 2012 Þ entrata in contatto con Hein van Grouw
(Life Sciences department - Natural History Museum - Tring - Hertfordshire - UK)
il quale si dedica alla ricerca relativa all'amelanosi negli uccelli, quell'amelanosi
che pu‗ fare la sua comparsa anche oltre le 20 settimane di vita, e magari
indipendentemente da fattori genetici. Hein van Grouw ha comunicato a Elly
quanto segue, che a mio avviso Þ di estremo interesse.
<<Progressive Greying, which may arise after a bird reaches a certain age and is defined as the progressive loss of pigment cells with age. From the onset of the condition, the bird will gain an increasing number of white feathers after every moult: finally the entire plumage becomes white. Progressive Greying may or may not be heritable. External, non-heritable factors such as illness or food deficiency can also be the cause of pigment loss. In such cases, the bird is unable to extract sufficient quantities of tyrosine from its food, which results in a disturbed melanin synthesis. Progressive Greying in an early stage shows white feathers spread randomly, often on the head, back and flanks at first, though often the skin (bill and feet) is unaffected.>>
<<Il Progressivo Ingrigire pu‗ insorgere dopo che un uccello ha raggiunto una certa etÓ, ed Þ definito come la perdita progressiva di cellule pigmentarie con l'etÓ. A partire dall'inizio del processo, dopo ogni muta l'uccello acquisirÓ un incremento del numero di penne bianche: alla fine l'intero piumaggio diventa bianco. Il Progressivo Ingrigire pu‗ essere o non essere ereditabile. Fattori esterni non ereditabili, come una malattia o la deficienza di cibo, possono anche essere la causa della perdita di pigmento. In tali casi l'uccello non Þ capace di ricavare quantitÓ sufficienti di tirosina dal suo cibo, il che dÓ luogo a una disturbata sintesi di melanina. In una fase precoce il Progressivo Ingrigire mostra delle penne bianche localizzate in modo irregolare, di solito prima di tutto su testa, schiena e fianchi, sebbene spesso la pelle (becco e piedi) non sia interessata.>>
L'importante ricerca di Hein van Grouw Þ stata pubblicata sulla rivista inglese British Birds (January 2013) nell'articolo intitolato <What colour is that bird? The causes and recognition of common colour aberrations in birds.>
La tirosina Þ un aminoacido biologicamente importante in quanto precursore di vari ormoni, quali la tiroxina (un ormone tiroideo) e le catecolamine (dopamina, noradrenalina e adrenalina), nonchÚ della melanina.
9. INCROCI TRA SOGGETTI BIANCHI E SOGGETTI DI ALTRO COLORE
Soggetto
bianco dominante x soggetto interamente nero
In F1
tutti i discendenti sono bianchi, con qualche possibile traccia di nero.
Reincrociando un soggetto di F1
con un bianco, oppure soggetti di F1
tra loro, otteniamo bianchi omozigoti (50% nel
primo caso, 25% nel secondo).
Soggetto
bianco recessivo x soggetto interamente nero
In F1
tutti i discendenti sono neri portatori del bianco. Incrociati tra loro
daranno: 25%
bianco
puro - 25% nero puro - 50% nero
portatore di bianco.
Soggetto
bianco dominante x soggetto dorato
In F1
il nero Þ giÓ sostituito dal bianco, come succede per il petto dei galli
dorati, mentre le parti rosse sono solo leggermente attenuate. Questo colore
Þ conosciuto come pile.
Incrociamo tra loro i pile
di F1: lÆazione di I
sul rosso si accentua perchÚ otteniamo soggetti omozigoti per I.
Le parti rosse saranno molto attenuate, da giallo ad arancio chiaro.
Per cui i pile incrociati tra loro danno dei discendenti
in cui il rosso scompare di generazione in generazione fino ad arrivare al
bianco puro, dato che I allo
stato omozigote cancella il nero e attenua molto il rosso. Quindi qualsiasi ceppo pile deve essere regolarmente
ritemprato con dei soggetti dorati. La stessa cosa si produce nel
Combattente indiano giubileo, il cui colore deve essere regolarmente ravvivato
attraverso incroci coi soggetti scuri.
| Maschio bianco dominante | Femmina nera | ||
|
I / I |
i+/i+ |
||
| I = bianco dominante i+ = allele di I |
|||
| 100%
soggetti bianchi con possibili tracce di nero |
gameti femminili |
||
| i+ | i+ | ||
|
gameti maschili |
I | i+/I | i+/I |
| I | i+/I | i+/I | |
| Maschio
bianco dominante eterozigote |
Femmina
bianco dominante eterozigote |
||
|
I / i+ |
I/i+ |
||
| I = bianco dominante i+ = allele di I |
|||
| 25% bianco
puro 50% bianco con tracce di nero 25% nero |
gameti femminili |
||
| I | i+ | ||
|
gameti maschili |
I | I/I | i+/I |
| i+ | I/i+ | i+/i+ | |
| Maschio bianco recessivo | Femmina nera | ||
|
c / c |
C+/C+ |
||
| c = bianco recessivo C+ = allele di c |
|||
| 100% soggetti neri |
gameti femminili |
||
| C+ | C+ | ||
|
gameti maschili |
c | C+/c | C+/c |
| c | C+/c | C+/c | |
| Maschio
nero eterozigote per c |
Femmina
nera eterozigote per c |
||
|
c / C+ |
c/C+ |
||
| c = bianco recessivo C+ = allele di c |
|||
| 25% bianco 50% nero portatore di c 25% nero puro |
gameti femminili |
||
| c | C+ | ||
|
gameti maschili |
c | c/c | C+/c |
| C+ | c/C+ | C+/C+ | |
10. I RIFLESSI PAGLIA
╚ giunto il momento di
analizzare una volta per tutte la vexata
qaestio dei riflessi paglia. Ricordiamo solo per inciso che taluni uccelli
conferiscono al proprio piumaggio bianco delle sfumature giallastre o rossicce
impregnandolo con la sostanza cerosa prodotta dallÆuropigio![]() : Þ cosý che
nasce la macchia gialla sul collo del Pellicano e la sfumatura rosata
posseduta da alcuni Gabbiani.
: Þ cosý che
nasce la macchia gialla sul collo del Pellicano e la sfumatura rosata
posseduta da alcuni Gabbiani.
Nel pollo i riflessi paglia sono di frequente riscontro e non esiste
allevatore che li veda di buon occhio. La stagione pi¨ propizia alla loro
comparsa - o alla loro intensificazione - Þ quella estiva. Una delle sedi
preferite Þ la mantellina dei galli purchÚ dotata di bianco, che non deve
necessariamente occupare unÆampia porzione di questo distretto. Ho potuto
spesso notare questi riflessi nei maschi di Amburgo pagliettata argento e di
Brahma bianco columbia. Nei soggetti completamente bianchi con piumaggio
soffice, oppure con piumaggio arricciato classificabile come ultrasoffice, i riflessi paglia possono invadere tutto il mantello e tutta quanta la
piuma. Molti allevatori concordano sul fatto che questi riflessi compaiono
pi¨ volentieri in soggetti bianchi non derivati dal nero esteso oppure
appartenenti al bianco recessivo.
10.1. Basi teoriche dei riflessi paglia
Esistono tre assiomi ricorrenti.
À I riflessi paglia del piumaggio bianco sono dovuti a residui di rosso nel mantello dei galli, nei quali sono spesso visibili. Dire che sono causati dal sole Þ falso, poichÚ i raggi attinici non fanno altro che metterli in risalto, essendo geneticamente determinati.
À
La sfumatura paglia Þ frequente nel bianco recessivo.
À Nei ceppi a bianco dominante i riflessi paglia sono possibili se la creazione del bianco Þ recente e debbono scomparire negli omozigoti dopo alcune generazioni.
Per il terzo assioma Þ quanto succede in pratica. In
teoria un bianco dominante omozigote neoformato dovrebbe avere le stesse
caratteristiche di quello che Þ frutto di alcune selezioni. Si pu‗ dedurre
che ce ne vogliono di incroci e di vittime per far scomparire il ricordo
genetico della feomelanina o di qualche altra diavoleria! Quale meccanismo
pu‗ essere invocato? Dobbiamo per forza chiedere soccorso ai soliti geni
modificatori, ai soliti ignoti
trascinati per i capelli dentro a un ceppo che Þ frutto di un lungo lavoro di
selezione?
Jeffrey Þ pi¨ sottile nellÆuso
della terminologia, in quanto distingue due tipi di colorazione anomala a
carico del piumaggio bianco:
À Riflessi color crema: sono evidenti a carico della totalitÓ della piuma, anche nellÆiporachide. La tendenza verso riflessi di questo tipo Þ ereditaria, ma la loro entitÓ pu‗ venir attenuata alimentando i soggetti con frumento e orzo anzichÚ granoturco e erba. Vorrei puntualizzare che una siffatta alimentazione ha un significato solo quando le piume stanno crescendo.
À Riflessi color ottone: sono limitati alla sola superficie esterna delle piume maschili e interessano la mantellina, il dorso, la sella e le piccole copritrici alari. LÆentitÓ di questi riflessi dipende dal grado di esposizione diretta alla luce solare. Questo difetto, come il precedente, Þ ereditabile e pu‗ essere ridotto con la selezione. Bisogna ricordare che Þ molto pi¨ difficile ottenere maschi bianco puro di quanto non avvenga con le femmine. Da questÆultima affermazione dovremmo arguire che potrebbero non essere estranei gli ormoni sessuali nel determinismo dei riflessi ottone.
Pi¨ sottile di Fred Jeffrey Þ
Eason Smith, le cui osservazioni sono riportate in The Australian Pekin Bantam. Secondo Smith bisogna distinguere:
À Brassiness o riflessi color ottone: si tratta di una colorazione congenita che si manifesta come sfumatura gialla a carico della mantellina e del dorso dei maschi, sia che vengano tenuti al riparo sia che vengano lasciati liberi di vivere una vita da pollo. Si tratta di un fattore limitato a un sesso: le sorelle di questi maschi sono portatrici del carattere e lo trasmettono alla prole. LÆunica soluzione per risolvere il problema Þ dedicarsi alla selezione, tenendo ben presente che non vanno assolutamente impiegate le sorelle di un soggetto che presenti questo difetto.
À Weathering o effetto da agenti atmosferici: si tratta di una colorazione marrone rossiccio, tannino, che viene acquisita vivendo allÆaria aperta. Se dopo la muta i soggetti vengono tenuti al riparo, il piumaggio si conserva perfettamente bianco.
In ambedue le suddette condizioni lÆalterazione
cromatica Þ limitata alla faccia esterna della parte distale della piuma,
cioÞ il vessillo, mentre sia il piumino che lÆiporachide rimangono bianchi.
À Sappiness: letteralmente abbondanza di linfa, succositÓ, sciocco in gergo USA. Questa situazione Þ presente nei giovani che hanno appena raggiunto la maturitÓ e si manifesta come sfumatura color crema che inizia dalla base della piuma per dirigersi verso lÆapice. Anche il colore del piumino Þ alterato come nel resto del piumaggio. Raggiunta la piena maturitÓ, abitualmente il color crema scompare del tutto dopo 1-2 settimane di vita allÆaperto, oppure in occasione del primo bagno in vista di una mostra. I soggetti con pelle e tarsi gialli sono generalmente pi¨ suscettibili a questÆanomalia transitoria rispetto ai soggetti a pelle bianca.
Come si pu‗ notare, Smith non accenna ai carotenoidi come
possibile causa dei riflessi paglia, salvo in modo indiretto, quando parla
della maggior incidenza della sappiness nei soggetti con cute gialla.
Da notare che la scomparsa della sappiness con la vita allÆaria aperta
coincide con quanto ho espresso a proposito dellÆeffetto scolorente sui
carotenoidi da parte dei raggi solari (vol.II - XXIX-1.)
e del fatto che la presenza o meno di carotenoidi nella piuma pu‗ essere
verificata con un bagno a base di acqua e sapone associato magari a qualche
solvente dei grassi. Un collegamento tra sappiness
e carotenoidi si pu‗ anche ravvisare nel consiglio di alcuni studiosi: essere
parsimoniosi in granoturco e erba per tutto il periodo antecedente la
maturitÓ sessuale.
Per dimostrare lÆeventuale
importanza dei carotenoidi nel determinare obbligatoriamente - da soli e nellÆetÓ
adulta - una colorazione gialla del piumaggio bianco, Þ stato intrapreso un
esperimento con la collaborazione di Eduardo Cani, partendo dallÆesperienza
dei canaricoltori che si dilettano nel colorare il Canarino con carotenoidi al
momento della muta
![]() -
- ![]() .
LÆesperimentucolo si Þ svolto cosý:
.
LÆesperimentucolo si Þ svolto cosý:
º maschio: Moroseta bianco con iride molto scura, becco ardesia depigmentato in punta
º femmina: ibrido usato per la cova, nana, forse dotata di bianco dominante, iride giallo chiaro, tarsi bianchi, becco chiaro.
╚ stato somministrato quotidianamente pane bagnato in
acqua con aggiunta di Red Factor,
cioÞ una miscela commerciale di carotenoidi e xantofilla
color mattone scuro usata
per i canarini. I pigmenti sono stati somministrati al mattino a partire dallÆ1-3-1995 al 15-5-1995
a dosi quotidiane di 0,5 g per la prima settimana e 1
g
per la seconda settimana. Non si sono osservate modificazioni apparenti della
pigmentazione dellÆocchio e dei tarsi della femmina; lÆocchio del maschio
Þ rimasto scuro come prima, soprattutto il piumaggio - piumino incluso - non ha
mostrato traccia di pigmento.
La durata dellÆesperimento non
Þ stata eccessiva. Tuttavia i carotenoidi avrebbero avuto tutto il tempo di
conferire almeno qualche nuance al
piumaggio, anche se debbo sottolineare che la coppia viveva a contatto dei
raggi solari. Che i carotenoidi passassero in circolo era dimostrato dallÆintensissima
colorazione del tuorlo. LÆesperimento verrÓ eventualmente ripreso in
occasione della prossima muta.
10.2. Il sebo-cheratinocita
Il sebo-cheratinocita del pollo
Þ stato ampiamente descritto nel II volume,
sezione XXVI - 1.3. alla quale si rimanda.
10.3. Risvolti pratici
Dotandoci di spirito dÆosservazione,
requisito fondamentale nelle argomentazioni scientifiche, dobbiamo ammettere
che in un pollo dal piumaggio compatto i riflessi paglia non interessano la
totalitÓ di una piuma appartenente alla mantellina: la piuma si presenta
giallognola solo in parte. Infatti le lanceolate sono embricate come le
tegole, per cui le piume superiori ricoprono parzialmente quelle poste pi¨ in
basso, e basta sollevare le lanceolate per rimanere colpiti dal candore delle
parti protette. Nel caso del piumaggio soffice oppure arricciato Þ facile
arguire che la luce riesce a penetrare praticamente sino al calamo, per cui Þ
facile che la sfumatura giallognola interessi tutta la piuma.
Una buona casalinga pu‗ insegnarci qualcosa circa le
interazioni tra luce e colore: Þ dÆobbligo che gli indumenti delicati
debbano asciugare al riparo dai raggi solari. Credo inoltre che tutti quanti
abbiamo avuto modo di intuire la labilitÓ delle tinte osservando lo
scolorimento delle foto esposte alla luce del sole per lunghi periodi di
tempo, sia essa diretta che indiretta. Per sbiancare
la biancheria, un tempo era
possibile stendere le lenzuola sullÆerba dÆun prato dopo averle bagnate,
insaponate e azzurrate col blu di metilene che oggi si trova sotto forma di
scagliette blu nei detersivi in polvere, ma che un tempo era confezionato in
palline, riunite poi dalla massaia in sacchettini da immergere nel bucato,
come si faceva anche per i blocchi e le scaglie di verderame. Posso
testimoniare che le lenzuola risultavano tanto candide da far annerire
le estenuanti pubblicitÓ televisive. E cosý gli allevatori pi¨ filoni non
risparmiano una passata dÆazzurro ai loro soggetti bianchi prima di portarli
in mostra!
Abbiamo quindi affermato che la luce solare possiede la
capacitÓ di scolorire, in particolare abbiamo giÓ detto che i carotenoidi
vengono affievoliti dalla luce e sappiamo inoltre che 72
ore
di esposizione sono in grado di alterare talmente il giallo da carotenoidi che
una piuma blu come quella dellÆAra ararauna ![]() diventa verde in quanto
viene a mancare il filtro giallo naturale. Alcuni autori negano la presenza di
carotenoidi nelle piume del pollo; solo Brandt e Willelms ne ammettono la
possibile presenza.
diventa verde in quanto
viene a mancare il filtro giallo naturale. Alcuni autori negano la presenza di
carotenoidi nelle piume del pollo; solo Brandt e Willelms ne ammettono la
possibile presenza.
Torniamo al giallo della mantellina. Se fosse dovuto ai
carotenoidi, tenderebbe a sbiadire, perchÚ scolorito dalla luce; inoltre il
giallo dovrebbe essere presente su una piuma da cima a fondo, e questo non
accade, anzi, Þ concentrato sulla parte scoperta della piuma e sulla faccia
esterna, mentre il lato rivolto verso la cute risulta bianco. Inoltre, se
fossero i carotenoidi a passare sulla piuma, incorreremmo in una palese
contraddizione, perchÚ in tal caso i riflessi paglia non farebbero la loro
comparsa su piume ancora in via di formazione (come ammesso da Crawford),
bensý su piume giÓ ultimate, magari prossime alla muta.
Non Þ concesso pensare che siano colpevoli i grassi dellÆuropigio![]() ,
in quanto il gallo dovrebbe superare Tiramolla in fatto di contorsionismo. Neppure noi riusciamo a
leccarci il collo e credo che neppure la giraffa riesca a farsi una toilette
completa in tale sede. Il pollo riesce a spulciarsi il petto e parte del
collo, ma nessuno avrÓ visto un soggetto capace di toccare col becco tutte
quelle piume che risultano cosý odiosamente gialle. LÆipotesi che il pollo
si passa sulle piume i carotenoidi dellÆuropigio come fa il pellicano (ma
pare che lÆuropigio del pollo non secerna carotenoidi) Þ unÆesagerazione,
in quanto il pollo non Þ un uccello acquatico e non ha perci‗ le necessitÓ
di un pellicano.
,
in quanto il gallo dovrebbe superare Tiramolla in fatto di contorsionismo. Neppure noi riusciamo a
leccarci il collo e credo che neppure la giraffa riesca a farsi una toilette
completa in tale sede. Il pollo riesce a spulciarsi il petto e parte del
collo, ma nessuno avrÓ visto un soggetto capace di toccare col becco tutte
quelle piume che risultano cosý odiosamente gialle. LÆipotesi che il pollo
si passa sulle piume i carotenoidi dellÆuropigio come fa il pellicano (ma
pare che lÆuropigio del pollo non secerna carotenoidi) Þ unÆesagerazione,
in quanto il pollo non Þ un uccello acquatico e non ha perci‗ le necessitÓ
di un pellicano.
Il mio Merlo Indiano ![]() , pur non essendo un acquatico, si spalma
accuratamente la secrezione dellÆuropigio su tutte le piume, ma questÆoperazione
quotidiana ha una giustificazione ovvia: se mi dimenticassi, ci pensa lui a
volare fino al lavandino e a richiedere con fischi acutissimi la ciotola di
acqua tiepida per le abluzioni alle quali non rinuncia affatto. Invece il
pollo, come altri Fasianidi, ama i bagni di sabbia o di terra. Lo stesso
discorso del prodotto di secrezione dellÆuropigio Þ valido anche per la
secrezione sebacea della cute del pollo, che nonostante tutto pare non
contenga carotenoidi.
, pur non essendo un acquatico, si spalma
accuratamente la secrezione dellÆuropigio su tutte le piume, ma questÆoperazione
quotidiana ha una giustificazione ovvia: se mi dimenticassi, ci pensa lui a
volare fino al lavandino e a richiedere con fischi acutissimi la ciotola di
acqua tiepida per le abluzioni alle quali non rinuncia affatto. Invece il
pollo, come altri Fasianidi, ama i bagni di sabbia o di terra. Lo stesso
discorso del prodotto di secrezione dellÆuropigio Þ valido anche per la
secrezione sebacea della cute del pollo, che nonostante tutto pare non
contenga carotenoidi.
Sappiamo che una piuma pu‗ colorarsi attivamente solo
mentre sta crescendo e che accade qualcosa di simile a ci‗ che si verifica al
tavolo della roulette: rien ne va plus,
les jeux sont faits. Abbiamo un bel dire che a passare nelle piume adulte
sono i carotenoidi del mais! Secondo questo principio saremmo costretti a
nutrire i nostri polli solo con erba! Ma neanche, in quanto proprio lÆerba
contiene i malefici pigmenti camuffati dalla clorofilla. Dovremmo quindi
ricorrere solamente a grano, orzo, mangime industriale privo di carotenoidi,
con la speranza di carpire almeno un sufficiente
alle competizioni.
Staples ha suggerito che i pigmenti che hanno ultimato la
loro deposizione su una piuma matura possono alterarsi per impregnazione della
corteccia della piuma da parte di una sostanza oleosa (che abbiamo visto
provenire dai sandwich epidermici).
Sý, ma possono alterarsi solamente i pigmenti. Ammesso che il bianco sia un
pigmento [orrore! Þ un colore, non un pigmento!], allora dovremmo ammettere
che nel pollo, il quale non usa spalmarsi in abbondanza di grassi, i
carotenoidi liposolubili passino per capillaritÓ dal derma a tutte le piume -
e in tutto il contesto della piuma - come accade per lÆuntuositÓ dei nostri
capelli (forniti di una o due ghiandole sebacee ciascuno), e come accade con
una goccia dÆolio che appunto sÆallarga
a macchia dÆolio su un foglio di carta. Se cosý fosse, lÆapice della
piuma dovrebbe perlomeno risultare meno impregnato rispetto alla base, come
dimostra la macchia sul foglio di carta, che presenta un alone periferico dove
lÆolio Þ meno concentrato rispetto al centro dove la goccia Þ caduta.
A questo punto dobbiamo fare un passo indietro, dando per
scontato il frequente
riscontro dei riflessi paglia in soggetti non derivati dal nero esteso.
Io reputo che nella genesi di un tipo di riflessi paglia succeda qualcosa di
simile a quanto accade per i capelli esposti al sole, i quali virano dal bruno
al biondo. Nei capelli la coesistenza di bianco e di nuances paglia non Þ incompatibile, anzi, Þ di frequente
osservazione
[8]
.
Si tratta di un ricordo di feomelanine? Il mio dubbio Þ il seguente: Þ
possibile che alcuni granuli di melanina, o meglio di feomelanina, non
visibili a occhio nudo ma presenti sulla piuma bianca, vengano trasformati
dalla luce solare fino ad assumere una colorazione paglia? Io reputo che
potrebbe essere vagamente possibile, visto oltretutto che il paglia
compare in aree colpite dalla luce. Sappiamo che nel Brahma bianco columbia la
base della piuma, lÆiporachide, pu‗ presentare una tonalitÓ grigioazzurra,
che invece deve essere assolutamente assente nella Sussex della stessa
varietÓ. Il gene del colore di base Þ differente per le due razze: nella
Brahma si tratta del perniciato, nella Sussex Þ in ballo il frumento. Ecco
quindi che due differenti geni possono condizionare la colorazione dellÆiporachide
in piume apparentemente identiche, e la nuance azzurrina a parer mio dovrebbe
essere dovuta a eumelanina.
Tuttavia, chi si appresta a esporre un pollo rovinato
da riflessi paglia, dovrebbe affrontare una prova molto semplice suggerita da
Eason Smith, ideata dal sottoscritto prima di conoscere i suoi suggerimenti e
mai realizzata: durante le abluzioni, praticate in
tandem
[9]
con il condizionamento del soggetto alla gabbia, si potrebbe lavare la
mantellina con abbondante sapone e con qualche solvente dei grassi, quali
trielina, benzina, etere, acetone. Io credo che il giallo non correlabile alla
sappiness
non scomparirebbe, perchÚ non Þ un giallo dovuto a carotenoidi disciolti nei
lipidi. Logicamente mi aspetto di essere presto confutato.
Secondo Martin esiste un metodo sicuro per far scomparire
i riflessi paglia dovuti a un residuo di rosso: utilizzare il gene dellÆargento, che cancella la
feomelanina.
In molte razze esistono le varietÓ argento e bianca. Quindi Þ molto facile
introdurre il gene S in un
ceppo bianco. Esistono delle Wyandotte il cui piumaggio ingiallisce e diventa
la disperazione dellÆallevatore, e altre invece il cui piumaggio Þ perfetto
perchÚ si Þ fatto ricorso ai consigli di Martin.
Sempre Martin ha fatto lÆesperienza con la Moroseta
incrociando grigio argento con bianco, che Þ un bianco recessivo. Lo scopo
era di migliorare la tipologia della grigio argento. Cosý, involontariamente,
in questo ceppo Þ stato incorporato un bianco recessivo eterozigote che fa
nascere regolarmente alcuni soggetti bianchi. I galli nati in questo modo non
ingialliscono mai.
Ciascuno ha i suoi consigli da elargire e vale la pena di
farne sempre tesoro. Cosý risparmieremo tempo e frustrazioni! Jeffrey consiglia di
creare il bianco miscelando il gene c
allo sfondo E, cui vale la
pena di aggiungere il blu. Il tocco finale Þ dato da S. Per cui la
formula del bianco perfetto Þ
la seguente: cc_EE_BlBl_SS.
Naturalmente, anche se Þ la formula della perfezione, Þ
una formula un poÆ lunga da realizzare, un poÆ come quelle ricette di
cucina che richiedono unÆintera giornata per un boccone prelibato. Ne
varrebbe tuttavia la pena. Esiste una scorciatoia per semplificare la
procedura: se lÆallevatore Þ angustiato dai riflessi paglia nei suoi
soggetti bianco recessivo, allora deve incrociarli con dei bei neri della
stessa razza. In prima generazione tutti i soggetti saranno neri. Si rimanda
all'ultimo quadrato di Punnett costruito ad
hoc per sapere cosa accadrÓ nelle generazioni successive, quando i
soggetti con genotipo E/E potranno
giÓ venir prescelti allÆatto della schiusa.
Non bisogna farsi ingannare dal piumino dei pulcini nel
lanciare pronostici sulla bellezza del bianco adulto. Senza dubbio i pulcini
colorati in rosso o in fulvo Þ meglio siano messi da parte, essendo
improbabile che rivestano un manto candido nella vita adulta. Parecchi pulcini
con un piumino fuligginoso oppure scuro posseggono spesso un bel bianco da
adulti. Alcuni ceppi di Plymouth Rock bianca, da bianco
recessivo associato al gene B, sono talmente scuri alla schiusa che si pu‗
persino dedicarsi alla separazione dei sessi in base alla chiazza occipitale
bianca. Il gene B del barrato legato al sesso non Þ stato introdotto solo nei
suddetti ceppi di Plymouth Rock; come abbiamo giÓ accennato a proposito del
gene I, il gene B Þ stato trovato anche in ceppi di Livorno bianca
particolarmente candidi dotati di bianco dominante, e ho la certezza che il
gene B Þ stato introdotto anche nel mio ceppo di Pekin bianca che sicuramente
Þ bianca a causa del bianco recessivo.
Gli allevatori si disperano non solo di fronte alle
Wyandotte bianche. Abbiamo visto che spesso sono angustiati dai riflessi paglia nei galli Brahma bianco columbia
e si chiedono con quali incroci possano farli scomparire. Stando alla formula
genetica, il bianco columbia della Brahma possiede il gene
del perniciato
eb responsabile del disegno primario.
I galli perniciati presentano un mantello dorato che Þ
pi¨ rosso di quello del Gallo rosso della giungla. Introducendo il gene Co,
il nero viene radunato nella mantellina, nelle remiganti, nella coda e nelle
piume dei tarsi. Per far scomparire il colore dorato Þ stato introdotto il
suo allele S, che tuttavia non
Þ dominante in modo
netto
su s+.
Se i due alleli si
trovassero associati, otterremmo un maschio arancio
columbia e non un bianco. Ma, dopo anni di selezione, si sarÓ pur
prodotto qualche gallo DOC
omozigote
per S, nel quale non pu‗ e
non deve comparire alcuna traccia di feomelanina. Invece questo non Þ
concesso. Le femmine di Brahma bianco columbia e di Amburgo pagliettata
argento, che hanno come gene di fondo
eb,
non presentano riflessi
paglia. I rispettivi maschi sý. Per cui ci troviamo di fronte alla brassiness,
secondo la classificazione di Smith.
Quali sono le ipotesi da azzardare e come deve essere
impostata una corretta classificazione dei vari tipi di riflessi paglia? A
questo punto sono molto disorientato in quanto la soluzione pu‗ venire solo
dallÆanalisi chimica delle sostanze responsabili dei diversi tipi di giallo.
Le femmine perniciate hanno lo stesso gene di fondo dei
maschi. PerchÚ non presentano la ripartizione delle melanine uguale a
quella dei maschi cosý da avere nei due sessi unÆidentica distribuzione dei
pigmenti, come
accade invece nella maggioranza dei colombi? Ancora nessuno ha dato una
spiegazione genetica molecolare a questo differente piumaggio del gallo e della
gallina.
La scopo dellÆappariscente piumaggio maschile Þ facile
da intuire: allo stato selvatico la femmina Þ mimetica in quanto spetta a lei
la cova e pertanto non deve e non pu‗ essere preda dei nemici, distolti dagli
smaglianti colori del gallo.
In certe razze create dallÆuomo, siano esse monocolori
che policrome, spesso la femmina ha il mantello dello stesso colore del gallo,
e nella Sebright lÆidentitÓ fra i sessi Þ spinta al massimo dal gene Hf
(vol.II - XV.5.1. - XXIV.4.)
che Þ in grado di assicurare un piumaggio monomorfo.
Per spiegare il dicromatismo sessuale nelle colorazioni di
base derivate da mutazioni di e+,
eccetto il nero
integrale, non basta affermare che la gallina possiede un solo gene s+
mentre il gallo Þ
omozigote per questo gene. Nella distribuzione e nella composizione della
feomelanina intervengono fattori da identificare. SenzÆaltro predominano gli
ormoni sessuali, come dimostrato dal cappone e dalla poularde, ma abbiamo
bisogno di lumi ulteriori.
Un esempio, ben visibile da lontano e che colpirebbe lÆosservatore
pi¨ disattento, Þ dato dalla colorazione frumento: non si direbbe affatto
che una frumentina abbia come partner un gallo molto simile al dorato. Che
dire poi del petto salmonato della femmina di Gallus gallus, mentre
quello del consorte Þ perfettamente nero! PerchÚ le femmine argentate con
genotipo e+ conservano il petto salmonato che non viene intaccato dal
gene S?
Sý, forse un solo gene argento Þ troppo poco per annullare un color salmone,
ma pur sempre salmone rimane. E non si tratta affatto di salmone sbiadito pur
essendo circondato dallÆargento! Solo il gene columbia associato allÆargento
pu‗ annullare il petto salmone.
LÆaffermazione che il gene S sia in grado di annullare gli effetti del suo allele nella
genesi dei riflessi paglia Þ senzÆaltro giusta quando sia
contemporaneamente presente il bianco recessivo, anchÆesso in grado di
inibire la feomelanina. Ma la stessa affermazione non potrebbe quadrare nel
caso in cui nessun gene del bianco Þ in ballo, come accade nel bianco
columbia e nel pagliettato argento, due disegni che non disdegnano la brassiness.
In
conclusione: o gli allevatori di polli caratterizzati da questi
disegni introducono nel bianco columbia anche il gene del bianco, e allora
addio nero; oppure debbono trovare o selezionare un ceppo in cui il ricordo
della feomelanina, o di altri pigmenti, sia diventato molto tenue. LÆinterazione
dei geni Þ complessa e la genetica non osserva la regola dellÆ1+1=2,
come abbiamo avuto modo di affermare a proposito del gene del blu.
11. MEDITAZIONE FINALE
Gran parte delle formule
genetiche possono sembrare difficili, e lo sono. Tuttavia non dobbiamo nutrire
unÆeccessiva repulsione nei loro confronti se vogliamo perfezionare i nostri
ceppi. Le formule ci fanno soprattutto capire che, pi¨ un disegno Þ
complesso, pi¨ un incrocio mal fatto Þ pericoloso, perchÚ in molti casi
pu‗ comportare delle combinazioni tanto strane da non riuscire a tener sotto
controllo la discendenza.
Se vogliamo avventurarci in colorazioni insolite, bisogna
essere certi della purezza dei genitori. Questa purezza Þ pi¨ rara di quanto
generalmente si creda: molti polli portano dei geni recessivi allo stato
eterozigote, quindi inattivi. Nel caso di un incrocio fatto a caso, o
lavorando in consanguineitÓ, tale gene pu‗ ritrovare il suo omologo e dare
un risultato tanto inatteso quanto indesiderabile.
Prendiamo come esempio il nero. Il nero integrale ha un
genotipo E/E senza alcuna
traccia di rosso. Combinato con I/I
dÓ un bianco perfetto. Ma essendo E
la pi¨ dominante delle mutazioni derivate da e+,
tutte le sue combinazioni con unÆaltra mutazione dello stesso locus daranno
un soggetto nero. Cosý tutti i polli E/ER,
E/eWh, E/e+, E/eb, E/ey, E/ebc sono neri in apparenza, ma tale
nero Þ impuro. Il piumino non sempre Þ scuro, e piume rosse possono
comparire volentieri nella mantellina del gallo. La discendenza si dissocerÓ,
per esempio, in soggetti neri puri E/E,
in altri impuri E/e+. Questi soggetti nero impuro,
utilizzati per creare un ceppo bianco, daranno dei discendenti bianchi, ma che
hanno forte tendenza a ingiallire.
Prendiamo lÆesempio della Moroseta nella varietÓ nera
che, anche se rara, talora la si incontra. Coloro che hanno provato a
incrociarla con la bianca sanno che si possono ottenere, insieme ai neri,
soggetti brunastri. Ci‗ Þ dovuto al fatto che si Þ incappati in un nero
impuro e al fatto che il bianco della Moroseta Þ un bianco recessivo. In
prima generazione Þ il gene parassita associato a E che si esprime nella metÓ dei soggetti non neri.
Una consanguineitÓ ben condotta Þ la miglior garanzia
contro lÆapporto di geni parassiti. Permette inoltre di avere una buona
certezza della purezza del proprio ceppo se nessun soggetto di colore anomalo
Þ apparso dopo alcuni anni di selezione.
La
genetica del pollo ha fatto dei grandi progressi ma ha dei limiti e alcuni
misteri restano ancora inspiegati. Cosý, perchÚ il ciuffo dellÆOlandese
nera ciuffo bianco Þ bianco, e non
subisce le modificazioni cromatiche del rimanente piumaggio? Qual Þ la formula completa
del colore quaglia?
La nozione di famiglie
di colori dovrebbe permettere di semplificare la discussione circa lÆomologazione
delle varietÓ. Prendiamo per esempio la famiglia E/E.
Quando un pollo esiste nel nero, Þ del tutto plausibile che esista anche
nelle varietÓ pomellata, cuc¨, blu o grigio perla.
I cambiamenti di famiglia debbono essere accolti con
sfiducia. Un buon esempio Þ la Nana di Giava, detta in inglese Rosecomb.
La varietÓ di base Þ il nero. La blu, la cuc¨ e la bianca sono ugualmente
ben tipizzate. Per ottenere la millefiori e la columbia Þ stato necessario
fare appello a incroci complessi e talora il tipo ne ha sofferto.
Il problema non Þ far apparire una cosiddetta varietÓ.
Tutti gli allevatori lo fanno un giorno o lÆaltro per caso, per errore, o
peggio, volontariamente. Ma queste cosiddette varietÓ sono quasi sempre
eterozigoti, cioÞ non hanno alcuna possibilitÓ di trasmettere fedelmente il
fenotipo alla discendenza.
Attualmente la quasi totalitÓ delle colorazioni omozigoti possibili Þ stata esplorata. Quel che resta da esaminare e da affrontare Þ il riuscire a fissare una varietÓ in una razza che non esiste ancora, come fece Lord Sebright.
Non Þ
facile e non bisogna affidarsi al caso.