Vol. 2° - VIII.
L’APPARATO DI GOLGI

Fig.
VIII, 1 - Camillo Golgi (Corteno, Brescia 1843 - Pavia 1926)
istologo e patologo
La scoperta di questo organulo risale al 1898 quando
Golgi, impregnando con acido osmico le cellule nervose del Purkinje, mise in
evidenza un reticolo perinucleare cui diede il nome di apparato reticolare interno.
Camillo Golgi nacque a Corteno, presso Brescia, il 7 luglio 1843, si laureň
in medicina a Pavia dove fece ritorno per dare lustro all’Istologia che l’aveva
sempre appassionato. Nel 1906 ricevette il premio Nobel per la medicina
insieme allo spagnolo Ramón y Cajal. Morě a Pavia il 21 gennaio 1926.
1. MORFOLOGIA dell’apparato di Golgi
1.1. Aspetto in microscopia ottica
Morfologia: č estremamente variabile da
una cellula all’altra e nella stessa cellula, in quanto puň cambiare con lo
stato funzionale. A volte si presenta come un reticolo denso di trabecole
anastomizzate, altre volte come una placca irregolarmente fenestrata, puň
assumere l’aspetto di sfere unite tra loro, potendo anche apparire in forma
anulare. Nelle cellule ghiandolari costituisce un reticolo molto denso. La
morfologia, nonostante sia variabile, č caratteristica di ciascun tipo
cellulare e sembra che la struttura dipenda dall’organizzazione della
cellula.
Dimensioni: sono variabili, molto grandi
nelle cellule ghiandolari e nervose, ridotte nelle cellule muscolari, e
cambiano con lo stato funzionale: molto sviluppate nelle cellule in
iperattivitŕ, poco sviluppate in cellule a riposo e ipoattive. Durante l’invecchiamento
cellulare l’apparato di Golgi diminuisce progressivamente fino a scomparire.
Localizzazione: č relativamente costante per
ogni tipo di cellula: in quelle di origine ectodermica l’apparato di Golgi
č polarizzato, situato fra il nucleo e la superficie dell’epitelio; anche
nelle cellule delle ghiandole esocrine si osserva una sua polarizzazione fra
nucleo e polo apicale o escretore; nelle ghiandole endocrine la posizione
occupata č variabile, salvo nella tiroide, dove l’apparato di Golgi si
trova in sede sovranucleare, fra il nucleo e il follicolo tiroideo dove viene
riversata la tireoglobulina.
1.2. Aspetto in microscopia elettronica
Al microscopio elettronico l’apparato
di Golgi si presenta come un organulo costituito da unitŕ elementari, le cisterne,
fenestrate e impilate, punto d’incontro della circolazione fra citomembrane.
Svolge un ruolo essenziale nel trasferimento e nell’imballaggio delle proteine prodotte dal reticolo endoplasmatico,
nella sintesi delle glicoproteine e dei mucopolisaccaridi. Dal punto di vista
ultrastrutturale nell’apparato di Golgi si riconoscono due livelli di
organizzazione: la cisterna e il dittiosoma.
·
La cisterna č l’unitŕ fondamentale del
dittiosoma, con forma di compartimento appiattito e fenestrato; č limitata da
membrane lisce; ha un diametro pari a 0,5¸1
mm.
·
Il dittiosoma
[1]
č un sistema lamellare formato per associazione e impilamento di piů
cisterne o sacculi; il numero di cisterne č variabile, in media da 5 a 8, ma
puň arrivare a 30 o anche piů; uno spazio di 10-15 nm separa ogni cisterna
da quella vicina (in questo spazio sono assenti i ribosomi e il glicogeno,
come anche nello ialoplasma che circonda ciascun dittiosoma). Ogni dittiosoma
č provvisto di due facce, fra le quali si trovano le cisterne impilate:
§
faccia
cis,
convessa, in rapporto col reticolo endoplasmatico e le vescicole di
transizione
§
faccia
trans,
concava, in rapporto con vescicole o vacuoli di secrezione
|
|
Fig.
VIII.
2 - Dittiosoma. Il
dittiosoma č il sistema lamellare che si trova raffigurato appena al di sopra
del REG. Le vescicole provenienti dal REG si fondono con la faccia immatura
del complesso di Golgi. Le proteine, contenute nelle vescicole, vengono
trasportate da altre vescicole alle cisterne, come mostrano le frecce. A un
certo punto le proteine vengono smistate e confezionate per essere inviate all’esterno
del dittiosoma: a destra sono raffigurati i granuli di secrezione, al centro
le proteine raggiungono la membrana plasmatica, a sinistra l’elaborato
raggiunge i lisosomi. |
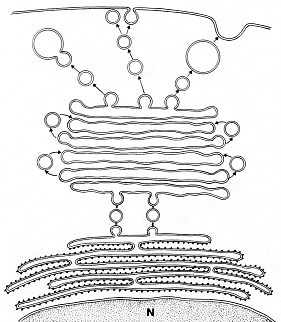
La faccia cis, le cui membrane
hanno uno spessore di 6 nm, paragonabile quindi a quello del reticolo
endoplasmatico, ha rapporti
con una cavitŕ del reticolo endoplasmatico granulare,
la cui membrana adiacente alla cisterna golgiana č priva di ribosomi. Questa
membrana dŕ origine, per
gemmazione, a piccole vescicole lisce, le vescicole di transizione,
con diametro di 20 nm. Vescicole piů voluminose, i granuli di secrezione, con
diametro compreso fra 40 e 80 nm, sono in rapporto con la faccia trans, le cui
membrane hanno uno spessore simile a quello della membrana plasmatica (7,5
nm).
Nonostante le numerose e approfondite ricerche sull’argomento,
dal punto di vista biochimico l’apparato di Golgi rimane ancor oggi un
grosso mistero. Infatti fino a qualche anno addietro si riteneva che tale
apparato fosse un sistema in continua trasformazione, nel quale una faccia immatura,
quella cis, si generava per apporto di vescicole provenienti dal REG, mentre
una faccia matura trans si consumava
per il distacco di macrovescicole e di vacuoli di secrezione diretti verso la
membrana plasmatica della cellula. Oggi si ritiene invece che l’apparato sia
costituito da cisterne relativamente stabili e compartimentate.
1.3. Funzionamento dei dittiosomi
Il traffico transgolgiano, secondo Farquar & Palade,
consiste in vescicole di transizione, formate per gemmazione del REL, che
trasportano prodotti qui elaborati. Le vescicole si fondono quindi coi bordi
dilatati delle cisterne golgiane. Il trasporto č pertanto assicurato da vettori vescicolari,
i quali si muovono, come navette,
fra il reticolo endoplasmatico e la periferia dell’apparato di Golgi. I
granuli di secrezione raggiungono la membrana citoplasmatica e, per un
processo di esocitosi, scaricano i loro prodotti di secrezione. Una delle
funzioni principali dell’apparato di Golgi č quella di smistare le proteine
di membrana, le proteine di secrezione e gli enzimi lisosomiali.
|
sommario |
avanti |
|
|
|