Vol. 1° - II.2.12.
Cosa serve per correre?
Per quanto Archaeopteryx non fosse probabilmente in grado di volare per lunghe distanze, era capace di effettuare il volo battuto ed era anche un buon corridore. In effetti la struttura del bacino e degli arti posteriori fa pensare che questo uccello preistorico fosse ben adattato a muoversi sul terreno. La conformazione del bacino - costituito da tre paia d'ossa: ileo, ischio e pube - ricorda quella dei dinosauri saurischi e soprattutto dei teropodi bipedi come Compsognathus. Anche i muscoli dell'addome e degli arti posteriori di Archaeopteryx, perciò, dovevano essere simili a quelli dei sauri.
È probabile che la posizione degli arti posteriori di Archaeopteryx, con il femore quasi verticale, fosse del tutto analoga a quella di Compsognathus e degli altri teropodi; viceversa, negli uccelli attuali il corpo è imperniato come un'altalena a livello del bacino e il femore è pressoché orizzontale. Come Compsognathus anche Archaeopteryx era bilanciato grazie al contrappeso della coda, che era lunga quasi quanto il corpo.
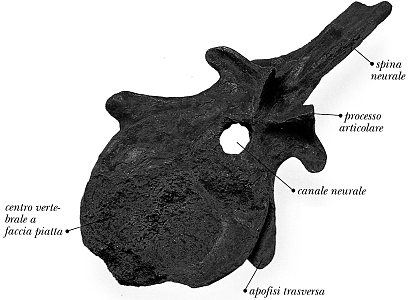
Fig. II. 10 - Vertebra di Cetiosaurus, appartenente all’Ordine dei Saurischi. Le sue vertebre a forma di chitarra sono molto caratteristiche, ma quello che a noi importa è il fatto che anche in Archaeopteryx il centro vertebrale - o corpo della vertebra - aveva superficie articolare quasi piatta.
La coda era flessibile alla base, ma diventava sempre più rigida verso l’estremità a causa delle sporgenze ossee delle 23 vertebre caudali [1] . Queste sporgenze sono presenti anche in alcuni dinosauri bipedi e nei sauri volanti dalla lunga coda che vissero nel Triassico e nel Giurassico. La rigidità della coda serviva all'animale per mantenere l’equilibrio in caso di improvvisi cambiamenti di direzione durante la corsa o il volo. Oltre a ciò, il rivestimento di penne della coda costituiva una superficie stabilizzatrice orizzontale.
Negli uccelli attuali le ultime vertebre della regione caudale sono ridotte e sono fuse insieme a formare il pigostilo - una delle poche strutture presenti solo negli uccelli - che sostiene le timoniere. Negli uccelli immediatamente successivi ad Archaeopteryx l’accorciamento della parte caudale della colonna vertebrale deve essere stato accompagnato da un graduale spostamento in avanti del baricentro. Per compensare in parte questo spostamento i muscoli della regione addominale aumentarono di dimensioni e vi fu un corrispondente incremento delle superfici d’inserzione dei muscoli sulle ossa del bacino. Nel corso della riorganizzazione del bacino le due ossa pubiche, dapprima fuse insieme, ruotarono all'indietro e si separarono. Il compito di sorreggere gli organi interni fu allora assunto dallo sterno, che contemporaneamente andò incontro a un notevole sviluppo.
La zampa di Archaeopteryx è senza dubbio ben adattata alla corsa e presenta caratteri intermedi fra quelli dei rettili e degli uccelli attuali. Nei rettili le ossa metatarsiche sono separate mentre negli uccelli attuali risultano fuse in un unico osso. L’esame ai raggi X dell’esemplare di Maxberg condotto da Wilhelm Stürmer - fisico e paleontologo alla Siemens di Erlangen - ha mostrato che le ossa del metatarso sono parzialmente fuse. Nell’esemplare più grande, quello di Solnhofen, le ossa appaiono fuse in misura ancora maggiore. In base a queste osservazioni si può dedurre che in Archaeopteryx le ossa del metatarso si ossificavano e si fondevano via via che l’animale cresceva.
Il piede di Archaeopteryx - e quello dei suoi antenati teropodi - è nel complesso simile per struttura a quello degli uccelli: presenta infatti tre lunghe dita e un quarto dito più corto rivolto all'indietro. L'artiglio acuminato e ricurvo del quarto dito fa pensare che questo uccello preistorico fosse in grado di afferrare oggetti con le zampe e di appollaiarsi sui rami degli alberi.
|
sommario |
avanti |
|
|
|